Бланк птс: «Бумажные бланки уничтожат». С 1 ноября ПТС стал электронным :: Autonews
ЛИЦЕВАЯ СТОРОНА ПАСПОРТА ТРАНСПОРТНОГО СРЕДСТВА (ОБРАЗЕЦ) / КонсультантПлюс
от 23 июня 2005 г. N 496/192/134
ПАСПОРТ ТРАНСПОРТНОГО СРЕДСТВА
00 АА 000000
Особые отметки Наименование (ф.и.о.) собственника _________ Особые отметки 1. Идентификационный номер (VIN) ___________
____________________________________________ ____________________________________________
Адрес ______________________________________ 2. Марка, модель ТС ________________________
____________________________________________ ____________________________________________
Дата продажи (передачи) ____________________ 3.
Документ на право собственности ____________ 4. Категория ТС (A, B, C, D, прицеп) _______
____________________________________________ 5. Год изготовления ТС _____________________
Подпись прежнего Подпись настоящего 6. Модель, N двигателя _____________________
собственника ______ собственника __________ 7. Шасси (рама) N __________________________
8. Кузов (кабина, прицеп) N ________________
М.П. М.П. 9. Цвет кузова (кабины, прицепа) ___________
10.
Свидетельство о регистрации ТС 11. Рабочий объем двигателя, куб. см _______
__________ серия _________ N _______________ 12. Тип двигателя __________________________
Государственный регистрационный знак _______ 13. Экологический класс ____________________
Дата регистрации ___________________________ 14. Разрешенная максимальная масса, кг _____
Выдано ГИБДД _______________________________ 15. Масса без нагрузки, кг _________________
____________________________________________ 16. Изготовитель ТС (страна) _______________
17. Одобрение типа ТС N _____ от ___________
Одобрение типа ТС N _____ от ___________
Отметка о снятии с учета ____________________________________________
Дата снятия с учета ________________________ 18. Страна вывоза ТС _______________________
М.П. Подпись _______________ 19. Серия, N ТД, ТПО _______________________
20. Таможенные ограничения _________________
Особые отметки Наименование (ф.и.о.) собственника _________ ____________________________________________
____________________________________________ ____________________________________________
Адрес ______________________________________ ____________________________________________
____________________________________________ 21. Наименование (ф.и.о.) собственника ТС __
Наименование (ф.и.о.) собственника ТС __
Дата продажи (передачи) ____________________ ____________________________________________
Документ на право собственности ____________ 22. Адрес __________________________________
____________________________________________ ____________________________________________
Подпись прежнего Подпись настоящего 23. Наименование организации, выдавшей
собственника ______ собственника __________ паспорт ____________________________________
____________________________________________
М.П. М.П. 24. Адрес __________________________________
Адрес __________________________________
____________________________________________
Свидетельство о регистрации ТС 25. Дата выдачи паспорта ___________________
__________ серия _________ N _______________
Государственный регистрационный знак _______ М.П. Подпись ______________
Дата регистрации ___________________________
Выдано ГИБДД _______________________________
____________________________________________
М.П. Подпись _______________
Отметка о снятии с учета
Дата снятия с учета ________________________
М. П. Подпись _______________
П. Подпись _______________
С 1 ноября 2019 года ПТС и свидетельство о регистрации на автомобиль будут выдаваться по новым формам
- Главная
- Правовые ресурсы
- «Горячие» документы
- Приказ МВД России от 23.04.2019 N 267 «Об утверждении форм документов, идентифицирующих транспортное средство, и требований к ним» (вместе с «Требованиями к паспорту транспортного средства», «Требованиями к свидетельству о регистрации транспортного средства»)
Приказ МВД России от 23.04.2019 N 267 «Об утверждении форм документов, идентифицирующих транспортное средство, и требований к ним» (вместе с «Требованиями к паспорту транспортного средства», «Требованиями к свидетельству о регистрации транспортного средства»)
Утверждение новых форм паспорта транспортного средства и свидетельства о регистрации транспортного средства связано с принятием Федерального закона от 03.08.2018 N 283-ФЗ «О государственной регистрации транспортных средств в Российской Федерации и о внесении изменений в отдельные законодательные акты Российской Федерации», вводящего в числе прочего возможность регистрации транспортных средств и получения госномеров непосредственно в салоне дилера.
Настоящим приказом установлено, что бланк ПТС должен изготавливаться на бумажной основе на листе размером 297 x 210 мм. ПТС, а также свидетельство о регистрации ТС будут иметь учетный номер и полиграфические защитные элементы. Оформление паспорта будет осуществляться путем заполнения его соответствующих строк и разделов с использованием принтера электронно-вычислительной машины (за исключением лазерного) или иного печатающего устройства.
В ПТС не допускаются приписки, подчистки, зачеркнутые слова и цифры, а также проставление прочерков. При ошибочном заполнении реквизитов изменения и дополнения вносятся в раздел «Особые отметки».
Свидетельство о регистрации автомобиля будет изготавливаться на бумаге размером 7,4 на 10,5 см, а затем ламинироваться. Данные о собственнике машины и название региона, в котором автомобиль состоит на учете, будут дублироваться на латинице. По желанию владельца транспортного средства записи в свидетельстве могут транслитерироваться в соответствии с данными, указанными в его загранпаспорте.
Приказ вступает в силу с 1 ноября 2019 года.
Перейти в текст документа »
Больше документов и разъяснений по коронавирусу и антикризисным мерам — в системе КонсультантПлюс.
Зарегистрируйся и получи пробный доступ
Дата публикации на сайте: 19.06.2019
Поделиться ссылкой:
В России вступили в силу изменения в водительских удостоверениях и ПТС
https://ria.ru/20201203/pts-1587449949.html
В России вступили в силу изменения в водительских удостоверениях и ПТС
В России вступили в силу изменения в водительских удостоверениях и ПТС — РИА Новости, 03.12.2020
В России вступили в силу изменения в водительских удостоверениях и ПТС
В России вступил в силу приказ министра внутренних дел Владимира Колокольцева, вносящий изменения в водительские удостоверения и паспорт транспортного средства. .. РИА Новости, 03.12.2020
.. РИА Новости, 03.12.2020
2020-12-03T08:41
2020-12-03T08:41
2020-12-03T13:08
общество
владимир колокольцев
министерство внутренних дел рф (мвд россии)
/html/head/meta[@name=’og:title’]/@content
/html/head/meta[@name=’og:description’]/@content
https://cdnn21.img.ria.ru/images/07e4/0c/03/1587491153_0:213:2826:1802_1920x0_80_0_0_00be06a014b966d21b764ad5c674a686.jpg
МОСКВА, 3 дек — РИА Новости. В России вступил в силу приказ министра внутренних дел Владимира Колокольцева, вносящий изменения в водительские удостоверения и паспорт транспортного средства (ПТС).Теперь в верхней части лицевой стороны прав будет располагаться надпись «Водительское удостоверение» на трех языках — русском, французском и английском.Кроме того, в ПТС будут вносить сведения о сроках государственной регистрации, количестве сидячих мест и технически допустимой массе автомобиля.Свидетельство о регистрации транспортного средства пополнится данными о мощности двигателя в лошадиных силах и таможенных ограничениях.
https://ria.ru/20201202/ekzamen-1587282847.html
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
2020
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
Новости
ru-RU
https://ria.ru/docs/about/copyright.html
https://xn--c1acbl2abdlkab1og.xn--p1ai/
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
https://cdnn21.img.ria.ru/images/07e4/0c/03/1587491153_0:0:2732:2048_1920x0_80_0_0_cc5123c46afbbbf6d51d39e35ad42ea2.jpgРИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
РИА Новости
internet-group@rian. ru
ru
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
общество, владимир колокольцев, министерство внутренних дел рф (мвд россии)
Новый бланк полиса ОСАГО и электронный ПТС
Пока все россияне заняты обсуждением повышения тарифов ЖКХ, начавших действовать с 1 июля 2016 года, а также вполне возможного дефицита заграничного алкоголя ввиду введения новых акцизов, то отечественные автолюбители в «мировой паутине» в это время рассматривают новый бланк полиса ОСАГО и гадают, чем обернётся для них переход от бумажных привычных ПТС к новым электронным.
Новый бланк страхового полиса ОСАГО
Начиная с 1 июля этого года, страховщики уже начали продавать страховые полисы на новых бланках ОСАГО. За всю историю этой страховки, впервые изменился цвет самого документа. Теперь он с зелёного сменился на розовый. Но это ещё не все новшества. Помимо этого, значительное внимание уделено увеличению степени защиты бланка. Эго новый фон стал довольно сложным, так как на нём появилась цветовая растяжка между розовым, жёлтым, и сиреневым цветами, а также графические фигуры.
Эго новый фон стал довольно сложным, так как на нём появилась цветовая растяжка между розовым, жёлтым, и сиреневым цветами, а также графические фигуры.
Дополнительно на бланке полиса ОСАГО проведено изменение шрифта, а также смена расположения и размера водяных знаков. Теперь на просвет по центру полиса видны изображения автомашины, а по бокам вертикально видно изображение логотипа Российского союза автостраховщиков. Кроме этого, в полис вставлена металлическая нить другого размера, располагающаяся также в другой части бумажного документа. На ней хорошо заметна надпись «ОСАГО», а на просвет ещё и надпись «полис». Также на новом бланке ОСАГО, в правом верхнем углу предусмотрено место для нанесения QR-кода. Первоначально, QR-код, ещё при отгрузке с Гознака, будет уже содержать в себе информацию о страховой компании, которой этот бланк был передан, ну, а впоследствии, при заключении договора страхования ОСАГО, в код будут внесены ещё и данные об автомобиле, его владельце и иная информация из в базы Российского союза автостраховщиков.
Представляя бланк нового образца, президент Российского союза автостраховщиков Игорь Юргенс, сообщил: «По информации Гознака, на изготовление подделки, мошенникам потребуется более двух лет только для того, чтобы суметь подделать металлизированную нить. Стоимость ОСАГО, при заключении договора страхования на новом бланке, не измениться и все дополнительные затраты «лягут на плечи» страховщиков, тем более, что они и так не велики. Стоимость изготовления бланка нового образца подросла меньше, чем на один рубль».
Будущее за электронным ПТС
В шести странах Евразийского экономического союза, включая и Российскую Федерацию, с 1 июля 2016 года вступило в действие соглашение о введении электронных паспортов транспортного средства (ПТС) для автомобилей.
Процесс перехода на электронный ПТС будет довольно длительным и растянется на два года, в течение которых органы ГИБДД будут продолжать параллельное оформление паспортов транспортного средства и на классическом бумажном носителе. Процесс постепенного перехода к новой форме основного документа на автомобиль будет длится до 1 июля 2017 года, ну, а для владельцев тракторов или иных самоходных транспортных средств процесс растянется аж до 1 июля 2018 года. В это время они пока также будут продолжать получать привычные бумажные ПТС. Вся информацию об авто в электронном виде планируется сохранять на серверах Компании «Электронный паспорт», входящей в структуру государственной «Ростехнология», которая утверждена, как единый администратор системы.
Но как, в своём интервью информагентству «Москва», сообщил первый зампредседателя комитета Госдумы по транспорту Михаил Брячак: «Паспорт транспортного средства — это совсем не единственный документ, который планируется в ближайшее время перевести в электронный вид. В России, возможно в скором времени, появятся ещё электронное водительское удостоверение.
TCP / IP Полная форма — javatpoint
TCP / IP: протокол управления передачей / Интернет-протокол
TCP / IP означает протокол управления передачей / Интернет-протокол. Это набор протоколов или правил и процедур, которые регулируют обмен данными между компьютерами в Интернете. Хотя весь набор интернет-протоколов обычно известен как TCP / IP, это один из основных протоколов Internet Protocol Suite. Он был разработан в 1978 году и управлялся Винтом Серфом и Бобом Каном.
Это набор протоколов или правил и процедур, которые регулируют обмен данными между компьютерами в Интернете. Хотя весь набор интернет-протоколов обычно известен как TCP / IP, это один из основных протоколов Internet Protocol Suite. Он был разработан в 1978 году и управлялся Винтом Серфом и Бобом Каном.
TCP / IP — широко используемый стандарт для передачи данных по сетям. Проще говоря, это набор протоколов связи, которые соединяют сетевые устройства в Интернете или используются для соединения сетевых устройств в Интернете. Он решает, как данные будут обмениваться через Интернет посредством сквозной связи, которая включает в себя то, как данные должны быть организованы в пакеты (пакеты информации), адресованы, отправлены и получены в месте назначения. Этот протокол связи также можно использовать для соединения сетевых устройств в частной сети, такой как интрасеть или экстранет.
Как работает TCP / IP?
Как следует из названия, TCP / IP состоит из двух основных протоколов: TCP (протокол управления передачей) и IP (Интернет-протокол).
TCP: TCP позволяет приложениям создавать каналы связи в сети. Это также позволяет разделить сообщение на более мелкие пакеты, прежде чем они будут переданы через Интернет, а затем правильно скомпилировать по адресу назначения. Таким образом, он обеспечивает надежную передачу данных по сети.Кроме того, он также проверяет ошибки в пакетах и запрашивает повторную передачу, если обнаружены ошибки.
IP: IP-адрес сообщает пакетам адрес и маршрут, чтобы они достигли нужного пункта назначения. У него есть метод, который позволяет шлюзовым компьютерам в сети, подключенной к Интернету, пересылать сообщение после проверки IPS-адреса. Это похоже на ряд рабочих, передающих уголь из шахты в карьерную тележку.
Уровни модели TCP / IP:
TCP / IP, который представляет собой стандартный многоуровневый набор протоколов, состоящий из набора правил и процедур, разделен на четыре уровня в зависимости от их функциональности.У каждого уровня есть определенный протокол.
- Прикладной уровень: Этот уровень включает в себя все протоколы, необходимые для прямой связи с конечными пользователями. Некоторые важные протоколы на этом уровне включают HTTP (протокол передачи гипертекста), FTP (протокол передачи файлов), SMTP (простой протокол передачи почты) и DHCP (протокол динамической конфигурации хоста).
- Транспортный уровень: Этот уровень обеспечивает передачу правильного сообщения или данных в правильном порядке.Он использует UDP (протокол дейтаграмм пользователя) и TCP.
- Уровень доступа к сети: Он предлагает функции для создания и обработки пакетов информации.
- Интернет-уровень: Он выполняет две основные функции: маршрутизацию и адресацию с использованием IP (Интернет-протокола). Он сообщает, как пакеты должны быть отправлены в пункт назначения.
Что означает TCP / IP? Что такое полная форма TCP / IP? »Английские аббревиатуры и акронимы» YThi
Полная форма TCP / IP — это протокол управления передачей / Интернет-протокол.
TCP / IP — это сокращение от протокола управления передачей / Интернет-протокола. TCP / IP часто рассматривается как набор согласованных протоколов, который был назван после объединения двух важных протоколов, а именно IP и TCP. Таким образом, его можно определить как основной элемент связи, применимый через Интернет.
Он представляет собой наборы сетевых протоколов, которые обеспечивают поддержку подключенных к сети компьютеров для легкого соединения с другими. TCP / IP, который является общеизвестным набором интернет-протоколов, состоит из наборов уровней, при этом каждый уровень отвечает за конкретную сетевую задачу и в то же время предлагает определенные услуги для верхнего уровня.Пакет Internet Protocol Suite был впервые разработан Агентством перспективных исследовательских проектов Министерства обороны США (DARPA) в 1970-х финансовых годах.
В принципе, TCP обычно применяется при передаче файлов данных и электронной почты. Протоколы нижнего уровня TCP / IP работают на смежных с пользователем уровнях и обрабатывают абстрактные данные. IP и TCP стали исходными протоколами, которые должны быть четко определены в стандарте Internet Protocol Suite.
IP и TCP стали исходными протоколами, которые должны быть четко определены в стандарте Internet Protocol Suite.
Именно в результате этих протоколов материализовалось имя набора протоколов.И наоборот, управление протоколом передачи функционирует в наборе интернет-протоколов, известном как транспортный уровень, где он предлагает подключенным к сети компьютерам надежные средства связи. В качестве сетевого протокола IP используется для передачи данных над сетью пакетных коммутаторов. IP облегчает передачу диаграмм, основанных на адресах сетевых IP-адресов, при работе на интернет-уровне набора протоколов.
В стопке протоколов TCP / IP протоколами самого высокого уровня являются протоколы приложений.Фактически, протоколы приложений являются видимым пользовательским интерфейсом для набора протоколов TPC / IP и всегда взаимодействуют с другими приложениями интернет-хостов. Каждый протокол приложения обладает функциями, аналогичными другим (Helmig n.p.). Характеристики, иллюстрирующие их использование и применение, следующие:
Протоколы TCP / IP — это приложения, которые транспортируются и стандартизируются с продуктами TCP / IP или приложениями, написанными пользователем. Набор протоколов для TCP / IP включает в себя протоколы приложений, а именно систему интернет-почты, называемую простыми протоколами передачи почты, и совместный терминал telnet, который обеспечивает доступ к удаленным интернет-узлам.Эти прикладные протоколы используют TCP или UDP в качестве транспортных механизмов. TCP предлагает ориентированные на соединение, надежные потоки, включенное управление потоком и протоколы, не допускающие перегрузок.
Набор протоколов для TCP / IP включает в себя протоколы приложений, а именно систему интернет-почты, называемую простыми протоколами передачи почты, и совместный терминал telnet, который обеспечивает доступ к удаленным интернет-узлам.Эти прикладные протоколы используют TCP или UDP в качестве транспортных механизмов. TCP предлагает ориентированные на соединение, надежные потоки, включенное управление потоком и протоколы, не допускающие перегрузок.
В качестве протокола одноранговой связи TCP не имеет ни подчиненных, ни главных корреляций. Тем не менее, для целей коммуникации приложение обычно использует модель сервер / клиент. Серверы — это приложения в TCP / IP, которые предоставляют потребителям Интернета необходимые услуги.
Пользователям нужно только вызвать сегмент клиентского приложения, который, в свою очередь, создает запросы на определенные услуги и отправляет их в сегмент сервера приложений через транспортное средство TCP / IP.Программа (сервер) принимает запросы, выполняет необходимые услуги и возвращает результаты в виде ответа.
Любой сервер в TCP / IP часто обрабатывает многогранные запросы клиентов и запросы одновременно (Panko 267). Таким образом, очевидно, что TCP / IP широко используется в моделях серверов и клиентов, а также для предоставления доступа к различным другим сетям через шлюз, мост и маршрутизатор.
| TCP | Протокол управления передачей Правительственные »Военные — и многое другое… | Оцените: | |||
| TCP | Tape Carrier Package Computing »Аппаратное обеспечение и многое другое … | Оцените: | |||
| TCP | The Children’s Place Бизнес »Компании и фирмы | Оцените: | |||
| TCP | Telesp Celular Participacoes Бизнес »Символы NYSE | Оцените: | |||
| TCP | Таба, Египет Региональные» Коды аэропортов | Оценить : | |||
| TCP | Центр инструмента Ac ademic & Science »Электроника | Оцените: | |||
| TCP | Торговая корпорация Пакистана Бизнес» Компании и фирмы | Оцените: | |||
| TCP | Программа управления терминалом Вычисления »Общие вычисления | Оцените: | |||
| TCP | Плазма с трансформаторной связью Академический и научный »Электроника | Оцените: | |||
| TCP | Протокол управления терминалом Правительственный» Военный | Оцените: | |||
| TCP | ThromboCytoPenia Медицина »Онкология | Оцените: | |||
| TCP | Точка контроля трафика Правительственный» Военный — и многое другое. | Оцените: | |||
| TCP | Простые люди Правительство »Правительство США | Оцените: | |||
| TCP | План управления транспортом Государственный транспорт | Оцените его: | |||
| TCP | Протокол управления передачей Вычислительная техника »Сети | Оцените: | |||
| TCP | Мощность телефонной связи Разное »Несекретный | Оцените: | |||
| TCP | Трансмиссия C ontrol Protocal Разное »Несекретный | Оцените его: | |||
| TCP | Надежная компьютерная платформа Вычислительная техника | Оценить это | |||
| TCP | Трикрезилфосфат Академия и наука »Химия | Оцените: | |||
| TCP | Тактический компьютерный процессор Государственный процессор | Оцените: | |||
| TCP | Процессор тактического управления Правительство »Военное дело | Оцените: | |||
| TCP 9007 3 | Программа по тропическим циклонам Академия и наука »Науки об океане | Оцените: | |||
| TCP | The Crowd Pleasers Разное» Несекретный 3 | Оцените: | |||
| TCP | Платформа сообщества учителей Сообщество »Образовательное | Оцените: | |||
| TCP | Передача Протокол контроля Разное »Несекретный | Оценить: |
 ..
..Официальный сайт LAWA | Транспортная чартерная партия
Добавить / удалить автомобиль
Каждый коммерческий автомобиль должен иметь действующее разрешение и активный транспондер, выданный LAX, прикрепленный к транспортному средству в порядке, установленном LAWA. Разрешения на автомобили и транспондеры не подлежат передаче или переуступке.
Разрешения на автомобили и транспондеры не подлежат передаче или переуступке.
Счета за все поездки в центральный терминал Лос-Анджелеса выставляются ежемесячно непосредственно Оператору по электронной почте в течение первых двух недель следующего месяца. Все счета подлежат оплате сразу после получения. Платежи можно удобно производить онлайн с помощью кредитной карты; используя ссылку, содержащуюся в вашем уведомлении о выставлении счета, или по почте через корпоративный чек или денежный перевод. Неуплата может привести к приостановке и / или прекращению действия NELA и права работать в LAX.Платежный портал GT Billing предназначен только для оплаты поездок.
Если у вас возникнут вопросы, пишите на [email protected]. Ваш адрес электронной почты должен включать название вашей компании, номер CPUC, контактное имя, номер телефона и причину вашего запроса.
Для добавления автомобиля:
- Заполните и отправьте форму добавления / удаления транспортного средства и копию действующей регистрации DMV коммерческого транспорта по адресу infolandside@lawa.
 org.
org.Документы должны быть одобрены персоналом GT, а оператор должен иметь:
- Активный статус CPUC
- Автомобиль должен быть на счете CPUC
- Утвержденная страховка и по расписанию транспортного средства
- Правильно отображаемые номера TCP на транспортном средстве, в соответствии с требованиями
- Все сборы уплачены; за каждый новый транспондер взимается плата в размере 50 долларов США
- Запишитесь на прием, чтобы получить новое разрешение и транспондер.
 org.
org.Назначить встречу
Для удаления автомобиля:
Заполните и отправьте форму добавления / удаления транспортного средства по адресу [email protected], верните транспондер и наклейку с разрешением на транспортное средство.
Разрешение на замену и / или транспондер
Лобовое стекло заменить? Назначьте встречу для замены транспондера, нажмите кнопку ниже. Принесите на прием старый транспондер и копию регистрации транспортного средства в DMV.
Назначить встречу
TCP против UDP — в чем разница между ними?
TCP и UDP — это оба способа передачи данных по сети, но, несмотря на то, что они одинаковы, они также разные.Вот что нужно знать об этих различных интернет-протоколах и о том, как они работают…
TCP vs UDP — в чем разница между TCP и UDP?
Когда дело доходит до понимания TCP и UDP — или того, что известно как протокол управления передачей и протокол пользовательских дейтаграмм , — нужно кое-что знать. В самом простом смысле все сводится к пониманию того, что они являются вариациями одного и того же и действуют по-разному.
Вкратце, это похожие (но разные) интернет-протоколы для передачи данных. В частности, TCP и UDP являются протоколами связи. Но это еще не все. Чтобы помочь вам лучше понять, мы расскажем, что такое TCP и UDP по отдельности — их сходство, их индивидуальные цели и принцип работы. После этого мы выделим разницу между TCP и UDP в удобной таблице.
После этого мы выделим разницу между TCP и UDP в удобной таблице.
Но сначала давайте лучше разберемся, что такое интернет-протокол, чтобы заложить основу для этой темы.
Что такое интернет-протокол?
Интернет-протокол (IP), тип сетевого протокола, представляет собой набор правил, которые определяют, как данные отправляются или передаются предполагаемому получателю через локальную сеть, через подсети или даже через Интернет. Так, например, всякий раз, когда вы отправляете данные из своего веб-браузера на веб-сайт — например, когда вы заказываете оптом лакомства для собак с вашего любимого веб-сайта с товарами для домашних животных, — вы делаете это через интернет-протокол.
Окольными путями передача данных по интернет-протоколу позволяет вам делать все, что вы делаете в сети — от твитов в социальных сетях и исследования рецептов до отправки электронных писем с изображениями или любимыми видео с кошками.По сути, это основа Интернета.
IP, TCP и UDP — все протоколы на уровнях модели OSI
Я упоминал, что IP — это сетевой протокол. Сеть — это лишь один из семи уровней модели взаимодействия открытых систем (OSI) ISO. Модель служит вертикальной структурой и стандартами, определяющими обмен информацией между компьютерными системами. Семь уровней модели OSI:
Сеть — это лишь один из семи уровней модели взаимодействия открытых систем (OSI) ISO. Модель служит вертикальной структурой и стандартами, определяющими обмен информацией между компьютерными системами. Семь уровней модели OSI:
- Физический уровень
- Уровень канала данных
- Сетевой уровень
- Транспортный уровень
- Сеансовый уровень
- Уровень представления
- Прикладной уровень
Обычно это визуализируется вертикально в порядке убывания, при этом физический уровень является самым нижним уровнем.В таком виде он традиционно выглядит примерно так:
Иллюстрация, которая описывает 7 уровней модели взаимодействия открытых систем ISO. С другой стороны, TCP и UDP — это два протокола, которые относятся к транспортному уровню модели OSI. На самом деле существует множество протоколов, которые подпадают под семь уровней и выполняют разные функции, и многие из них построены друг на друге. Сегодня мы не будем вдаваться во все это, поскольку цель этой статьи — помочь вам разобраться в TCP и UDP.
Прежде чем мы перейдем к подробному обсуждению разницы между TCP и UDP, вам сначала необходимо понять, как работает Интернет-протокол. Я говорю это, потому что TCP и UDP полагаются на IP. Это послужит основой для объяснения двух других концепций.
Как работает IP для передачи данных
Независимо от того, что вы отправляете, данные передаются взад и вперед по Интернету посредством процесса, известного как маршрутизация .Как вы, наверное, догадались, этот процесс включает использование маршрутизаторов или шлюзов для регулирования трафика. Согласно статье TechRepublic сетевого инженера и технического писателя Брайена Поузи, маршрутизаторы обычно предназначены для аналогичных сетей, тогда как шлюзы помогают соединять разнородные сети.
Теперь, чтобы гарантировать, что данные достигают предполагаемого получателя, каждому отдельному устройству назначен собственный IP-адрес — так же, как у вас есть конкретный адрес для вашего дома или квартиры. Но вместо 123 Main Street ваш IP-адрес в Интернете может выглядеть примерно как 250.250.250.250 (если вы используете стандартный адрес IPv4). Этот 32-битный номер состоит из серии подсетей (сегментов чисел, разделенных точками), каждая из которых содержит до трех цифр (до 12 цифр или 15 символов, включая точки).
Конечно, это выглядит иначе для IP-адресов IPv6, которые представляют собой 128-битные числа и выглядят примерно так: 2001: 0db8: 85a3: 0000: 0000: 8a2e: 0370: 7334 (это пример IPv6-адреса из Википедии).
Перед передачей ваших данных IP разбивает их на более мелкие PDU — они известны как пакеты , (для TCP) или дейтаграммы , (для UDP). Пакеты — это наиболее часто используемый термин для обозначения единиц данных в сети. Такое разбиение данных происходит при любой передаче данных IP — как TCP, так и UDP. Каждый из этих небольших пакетов данных снабжен меткой (, заголовок, ), которая сообщает доставщику, куда он должен идти и откуда он был отправлен.
Данные передаются маршрутизаторам или шлюзам , которые имеют списки информации для различных IP-адресов (и их соответствующих доменов) через Интернет. Информация передается от одного шлюза к другому, пока один из них не распознает адресат пакета или дейтаграммы.
Теперь просто имейте в виду, что это (очень) упрощенная разбивка того, как работает IP. Просто запомните эту иллюстрацию, потому что я вернусь к ней позже и применим TCP и UDP в качестве методов доставки.
Если вам интересно, какое место во всем этом занимает система доменных имен (DNS) (поскольку это, по сути, то, что связывает домены с соответствующими IP-адресами), просто знайте, что это взаимосвязано, но отдельно. Например, система доменных имен может использовать UDP для DNS-запросов. Однако мы не собираемся здесь подробно останавливаться на этом, потому что это всего лишь отдельный процесс, и в этой статье основное внимание уделяется различиям между TCP и UDP. Итак, в будущей статье нам придется более подробно рассказать о системе DNS.
Итак, в будущей статье нам придется более подробно рассказать о системе DNS.
Интернет-протокол — это служба доставки Интернета
Хорошо, давайте рассмотрим все это в небольшом контексте. Подумайте об интернет-протоколе, таком как FedEx, DHS или USPS в Интернете, но с небольшим поворотом (я объясню позже). С помощью этих типов служб доставки почты вы можете выбрать отправку посылок с помощью базовой доставки или попросить кого-нибудь расписаться за посылками по прибытии, чтобы убедиться, что они были доставлены нужному человеку. Интернет-протокол работает примерно так же, но для передачи цифровых данных.
Теперь, продолжая аналогию со службой доставки, представьте, что ваша мама хочет отправить вам ПУЧКУ домашней выпечки: печенье, хлеб, сушеные фрукты — вы называете это. (Тебе повезло, дьявол.) Для этого ей понадобится ваш адрес, чтобы негабаритная посылка была успешно доставлена к вашему порогу. В противном случае эти вкусные угощения могут оказаться в руках ваших нелюбимых соседей, Грега и Джилл (а это никому не нужно).
Итак, в этом случае ваша мама все упакует, пометит посылку вашим адресом ( заголовок ) и попросит ее любимую службу доставки приехать за ними.(Опять же, я , на самом деле , упрощаю здесь — позвольте мне немного расслабиться.) Однако ей приходится разбивать коробку на серию помеченных меньших коробок ( пакетов или дейтаграмм ), потому что это слишком многое нужно обработать.
Как только пакеты покидают ее дом, они попадают на станцию пересылки (, шлюз, ), чтобы добраться до следующего пункта доставки, ближайшего к месту назначения. Там пакеты продвигаются от станции к станции (от шлюза до шлюза ), пока не доберутся до той, которая знает ваш точный адрес.
А теперь представьте, что этот вкусный пакет — это данные, которые передаются через Интернет. Теперь, что происходит с этими пакетами в этот момент, будет зависеть от того, происходит ли доставка через TCP или UDP. Я сейчас объясню, что я имею в виду, но сначала давайте разберемся, что такое TCP и UDP и что влечет за собой каждый метод передачи данных.
Я сейчас объясню, что я имею в виду, но сначала давайте разберемся, что такое TCP и UDP и что влечет за собой каждый метод передачи данных.
TCP против UDP: все, что вам нужно знать об этих протоколах
Что такое TCP?
TCP означает протокол управления передачей , который является типом метода передачи данных с установлением соединения .Он был разработан, чтобы помочь установить надежные соединения практически в любой сети, даже в ненадежной. TCP, зависящий от IP, является частью транспортного уровня модели взаимодействия открытых систем (OSI) ISO. Это означает, что это протокол связи транспортного уровня.
Но как узнать, что вы используете TCP? Вы знаете этот символ буферизации, который иногда появляется, когда вы пытаетесь загрузить высококачественное видео на свой любимый видео-сайт? Часто это признак того, что данные, которые вы получаете, передаются через TCP.
По сути, цель TCP — быть безошибочным методом передачи данных. Протокол управления передачей помогает:
Протокол управления передачей помогает:
- Убедитесь, что получатель получает все отправленных данных (через квитанцию о доставке), и
- Сохраняйте передачу данных организованной, обеспечивая получение пакетов получателем в предполагаемом порядке.
TCP делает это, устанавливая надежное соединение, которое помогает ему проверять успешность получения пакетов.Обычно, если данные поступают так, как задумано, в том порядке, в котором они должны поступать, они получают подтверждение от получателя. В противном случае TCP заставит отправителя повторно отправить данные.
Протокол TCP был впервые определен в 1980 году Инженерной группой Интернета (IETF) в своем RFC 761. Позже он был отменен RFC 793 в 1981 году. Исторически это был очень распространенный метод передачи данных — тот, который вы, вероятно, используете каждый день, когда вы проводите свой день в Интернете.
Однако он не идеален и не может быть самым подходящим протоколом для использования во всех случаях. Именно здесь в игру вступает UDP (или другие протоколы связи). (Подробнее об UDP.)
Именно здесь в игру вступает UDP (или другие протоколы связи). (Подробнее об UDP.)
Как работает TCP при обмене данными
Протокол управления передачей требует установления соединения посредством трехстороннего квитирования перед передачей пакетов данных от сервера к клиенту. Трехэтапный процесс установления связи, который начинается с обмена сообщениями SYN, SYN-ACK и ACK (известными как флаги) между этими объектами, помогает обеспечить надежное соединение между ними.Выглядит это примерно так:
Иллюстрация трехстороннего рукопожатияТри сообщения следующие:
- SYN — синхронизировать
- SYN-ACK — синхронизировать-подтверждение
- ACK — подтвердить
Конечно, есть ряд других флагов, которыми можно обмениваться после этого момента, но мы не будем разобраться во всем этом сейчас. Давайте продолжим с целью процесса. После того, как это соединение установлено, каждый сегмент данных, который отправляется от клиента на сервер, сопровождается флагом ACK, подтверждающим получение пакета данных.
Теоретически все это звучит великолепно. Но какой реальный пример TCP в действии вы можете увидеть? По состоянию на июнь 2016 года TCP использовался сервисом потокового интернет-видео Netflix. (Однако я не уверен, продолжают ли они это делать или сейчас используют другой протокол, поскольку они так и не ответили на мой запрос для прессы.)
Что такое UDP?
Как мы упоминали ранее, UDP означает протокол пользовательских дейтаграмм . Хотя это похоже на TCP в том смысле, что это тип протокола связи транспортного уровня, он отличается, потому что это протокол связи без установления соединения — это означает, что соединение не должно устанавливаться перед отправкой ваших пакетов данных (или, более конкретно, дейтаграмм). .
При использовании UDP трехстороннее квитирование не требуется перед передачей данных предполагаемому получателю. Таким образом, все или некоторые данные могут прибыть — а может прибыть (а может и не) прибыть в том порядке, в котором он был запланирован. По сути, это часто называют «максимальным усилием».
По сути, это часто называют «максимальным усилием».
Существует отличная аналогия, которую пользователь форума stackexchange.com по имени Майк Робинсон использовал для описания протокола пользовательских дейтаграмм:
UDP помещает сообщение в бутылку и бросает его в море.Он может никогда не прийти. Он может прибыть до или после другой бутылки. Вы никогда не узнаете, прибыл ли он. Вы даже не знаете, жив ли вообще получатель, не говоря уже о том, чтобы вытаскивать бутылки из воды. Единственный способ узнать, было ли ваше сообщение услышано, — это если на вашу береговую линию вымыта еще одна бутылка, и вам удастся схватить ее до того, как она затонет ».
UDP, как и TCP, существует уже давно. Концепция протокола пользовательских дейтаграмм была впервые определена в IETF RFC 768 в 1980 году.В то время как TCP долгое время считался основным протоколом, UDP набирает обороты, поскольку скорость и производительность сети становятся приоритетом.
Как UDP работает при обмене данными
Преимущество UDP над TCP состоит в том, что он предлагает меньшую задержку, но загвоздка в том, что это также может привести к потере некоторых данных и их целостности. И, в отличие от TCP, если некоторые из этих данных отсутствуют, отправитель не отправит их повторно, потому что у них нет возможности следить за тем, чтобы вы все получили.
Все это говорит о том, что UDP не является безошибочным способом обмена данными. Таким образом, хотя это более быстрый процесс передачи данных, чем TCP, предостережение заключается в том, что вы не получаете данные точно так, как они были отправлены изначально, и данные могут отсутствовать (и вы никогда не узнаете об этом).
UDP идеально подходит для «живых» ситуаций, когда передача данных должна происходить в реальном времени без задержек. Некоторые возможные примеры использования UDP включают онлайн-игры в видеоигры и аудио- или видеозвонки.
Краткое описание различий между TCP и UDP
По сути, и TCP, и UDP являются протоколами, которые помогают определить, как данные обмениваются между двумя объектами. Самая большая разница между методами доставки TCP и UDP сводится к:
Самая большая разница между методами доставки TCP и UDP сводится к:
- Если и как подключаются. TCP — это все о соединении. Это протокол, который устанавливает надежные соединения — даже в ненадежных сетях — перед передачей данных. UDP — это протокол без установления соединения, что означает, что он будет отправлять данные независимо от того, может ли он подтвердить, что получатель находится здесь, чтобы получить их.
- Полнота и достоверность данных. TCP отдает приоритет целостности данных и гарантированной доставке, тогда как UDP отдает приоритет скорости за счет потери данных.
- Скорость передачи данных. TCP — претендент в тяжелом весе, тогда как UDP — чемпион в легком весе. В первом есть ряд процессов, которым необходимо следовать, чтобы обеспечить хорошее выполнение работы, а во втором — все для быстрого выполнения работы.
- Приложения для каждого протокола. TCP лучше всего подходит для доставки потоков данных более высокого качества, которые могут буферизоваться (например, сервисы потокового видео), тогда как UDP лучше всего подходит для приложений передачи данных в реальном времени, таких как аудио- и видеочаты, онлайн-игры, запросы DNS и т. Д.
 Д.
Д.TCP против UDP: Различия в сроках доставки данных (пример)
Давайте вспомним аналогию с домашней выпечкой вашей мамы и применим TCP и UDP в качестве методов доставки. Когда компания по доставке перемещает пакеты с выпечкой, пакеты будут доставлены по разным маршрутам до места назначения.Они могут или не могут прибыть в предполагаемом порядке или в том состоянии, в котором они были отправлены, и их могут просто высадить без подтверждения того, что вы находитесь там, чтобы их получить.
С TCP прибывают меньшие пакеты данных (выпечка), и это похоже на то, что есть личный помощник по доставке, который собирает их обратно в точном порядке, в котором они были отправлены вам вашей мамой. Таким образом, в оригинальной упаковке ничего не изменилось и не пропало — и вы должны расписаться за посылку по прибытии. Таким образом, ваша мама может гарантировать, что вы получите свои лакомства.Если кто-то заблудится, она узнает об этом и попытается отправить его повторно.
С другой стороны, с UDP дейтаграммы (запеченные элементы) могут быть изменены или немного переставлены с точки зрения порядка. А когда они прибывают в пункт назначения, их просто бросают у вас на пороге — им никто не подписывает, и никакая служба не собирает все заново для вас. Это означает, что если вы получите только часть пакетов с выпечкой, это очень плохо, потому что ваша мама не знает и не собирается повторно отправлять недостающие. Кроме того, поскольку у вас есть целая куча пакетов, вы даже не заметите, что один или два могут отсутствовать.
TCP против приложений UDP: когда один метод лучше другого?
Если вы хотите надежную передачу данных, TCP для вас. TCP гарантирует, что вы получите все точно в том виде, в каком оно было отправлено, и вам все равно, может быть задержка или отказ в соединении, и вам придется повторить попытку, тогда вы захотите использовать TCP. Но если вас больше волнует скорость и вы хотите получать информацию быстрее, независимо от того, была ли она получена точно так, как была отправлена, или если пакеты данных отсутствуют, тогда UDP работает нормально.
По сути, выбор между двумя протоколами сводится к тому, чего вы пытаетесь достичь. Некоторые примеры того, когда UDP будет предпочтительнее TCP, включают использование видео- и аудиочатов в реальном времени в Интернете, а также многопользовательские игры или поля битвы в живых онлайн-играх.
TD; DR — Ключевые различия между TCP и UDP
Послушайте, я понял. Вы заняты, и вам нужно много узнать о TCP и UDP. Оба эти протокола имеют свои преимущества и служат конкретным целям в нашем цифровом мире.Итак, чтобы быстро подвести итог для тех из вас, кто спешит, вот таблица, в которой представлены различия между TCP и UDP.
| TCP (протокол управления передачей) | UDP (протокол дейтаграмм пользователя) |
| TCP — это протокол связи с установлением соединения. | UDP — это протокол связи без установления соединения. |
Единицы данных TCP известны как пакеты. | Блоки данных UDP обычно называют дейтаграммами (хотя иногда их также называют пакетами). |
| TCP переупорядочивает пакеты данных для получения в предполагаемом порядке. | UDP отправляет дейтаграммы независимо, что означает, что они могут поступать в другом порядке. |
| Использует трехстороннее рукопожатие для установления надежных соединений. | UDP не использует процесс подтверждения. |
| TCP подтверждает получение данных о доставке. | UDP не использует квитанции о доставке. |
| TCP приводит к увеличению задержки. | UDP разработан для более быстрой передачи данных. |
| TCP гарантирует доставку данных, отдавая приоритет целостности, полноте и надежности данных. | UDP отдает приоритет скорости и часто приводит к потере данных. |
| TCP идеально подходит для надежной передачи данных. | UDP идеально подходит для передачи данных в реальном времени и потоковой передачи, чтобы избежать задержек. |
2021 Форма регистрации волонтера TCP
Virtual Job Shadow SponsorВиртуальная слежка за работой — это форма экспериментального обучения, предназначенная для предоставления студентам краткого практического опыта в интересующей их области.Принимая у себя студента на день, у вас будет возможность принять участие в исследовании его карьеры и процессе принятия решений, а также у вас будет возможность выявить потенциальных будущих стажеров, учеников или новых сотрудников. Студенты получают непосредственный опыт работы в технической профессиональной среде.
Демонстрации STEM
Волонтерство для руководства демонстрациями STEAM повышает грамотность школьников по STEAM и способствует развитию энтузиазма в отношении проектов в области науки, технологий, инженерии, искусства и математики.Волонтеры ищут STEM-проект, который они хотели бы провести через нашего партнера по программе, выражают заинтересованность в поддержке деятельности, а затем связываются с нашим офисом в Интернете для доставки и демонстрационных мероприятий.
Молодежная программа «Спросите эксперта»
Эта возможность дает экспертам возможность работать вместе над решением проблем, которые могут возникнуть у учащихся и / или родителей на любом этапе проекта STEAM. Добровольцы приезжают в назначенный день один раз в неделю через Zoom в течение 30 минут, чтобы ответить на вопросы, соответствующие их области знаний.Добровольцы должны иметь возможность уделять не менее 30 минут каждую неделю в запланированный день и время до конца мая.
Послы выпускников
Послы выпускников взаимодействуют с перспективными студентами, руководством TCP и, что наиболее важно, однокурсниками, чтобы продвигать захватывающие программы, разработанные для связи студентов с выпускниками, помогая студентам, когда они начинают свой путь в технологической карьере Программируйте и отмечайте, что значит быть выпускником TCP.
Корпоративный чемпион
Корпоративный чемпион рассказывает о своем опыте волонтерства TCP в своей компании и / или организации и вдохновляет других на участие. TechBridge предоставляет вам колоду волонтеров, темы для обсуждения и рекламные материалы на одной странице, которыми вы можете поделиться с отделами или отдельными лицами.
TechBridge предоставляет вам колоду волонтеров, темы для обсуждения и рекламные материалы на одной странице, которыми вы можете поделиться с отделами или отдельными лицами.
Success Coach Mentor
Успех коучей оказывает сильное положительное влияние на студентов в различных личных, академических и профессиональных ситуациях.Роли наставника по успеху открыты для профессионалов ИТ-индустрии, которые имеют опыт профессиональной работы не менее 3-5 лет и могут взять на себя долгосрочное (9-12 месяцев) обязательство помочь студентам изучить карьерные устремления, постановку целей, реализацию и оценку стратегий. и делиться мудростью и советами, которые приводят к погружению в технологическую экосистему, личностному росту и повышению самооценки.
Спикер-волонтер / участник дискуссии
Спикеры / участники дискуссии — это профессионалы и преподаватели, которые могут дать высококачественное финансовое образование, профессиональное развитие, получить представление о технических отраслях или выступить в качестве участников дискуссии для широкого спектра семинаров и / или презентаций в Программа технологической карьеры.
Границы | Факторы транскрипции TCP на границе между экологическими проблемами и реакцией роста растений
Обнаружение TCP —
Пелория и другие мутантыПластичность развития важна для выживания растений, потому что растения — это сидячие организмы, которые должны приспосабливаться к неоптимальным условиям окружающей среды. Крайне важно, чтобы эти адаптационные процессы развития были сбалансированы, а это означает, что необходимо воспринимать и взвешивать несколько стимулов окружающей среды, прежде чем растение наладит свой рост.Следовательно, множество регуляторных белков участвует в управлении реакциями развития на окружающую среду. Одно семейство факторов транскрипции, которое участвует во множественных процессах развития, — это белки TEOSINTE BRANCHED 1, CYCLOIDEA, PCF1 (TCP).
Лен обыкновенный ( Linaria vulgaris ) — многолетнее растение с двухсторонними зигоморфными цветками, произрастающее в Европе и на значительной части северной Азии. Когда Карлу Линнею подарили лен обыкновенный, у которого не были зигоморфные, а радиально-симметричные цветки, он назвал его peloria от древнегреческого πέλωρ (пелор), что означает монстр .Линней предположил, что это чудовище было гибридом льна обыкновенного и неизвестного до сих пор растения, и был удивлен, увидев, что этот гибрид, тем не менее, мог размножаться через семена (Linnaeus and Rudberg, 1744). В то время как его гибридная гипотеза оказалась ошибочной, он использовал этот случай как доказательство против неизменности, веры в то, что все виды созданы в начале мира и неизменны (Smith, 1821). Peloria — естественная разновидность льна-жабы, львиного зева ( Antirrhinum majus ) (Darwin, 1868) и наперстянки ( Digitalis purpurea ) (Keeble et al., 1910), среди других видов.
Когда Карлу Линнею подарили лен обыкновенный, у которого не были зигоморфные, а радиально-симметричные цветки, он назвал его peloria от древнегреческого πέλωρ (пелор), что означает монстр .Линней предположил, что это чудовище было гибридом льна обыкновенного и неизвестного до сих пор растения, и был удивлен, увидев, что этот гибрид, тем не менее, мог размножаться через семена (Linnaeus and Rudberg, 1744). В то время как его гибридная гипотеза оказалась ошибочной, он использовал этот случай как доказательство против неизменности, веры в то, что все виды созданы в начале мира и неизменны (Smith, 1821). Peloria — естественная разновидность льна-жабы, львиного зева ( Antirrhinum majus ) (Darwin, 1868) и наперстянки ( Digitalis purpurea ) (Keeble et al., 1910), среди других видов.
Примерно 250 лет спустя Луо и др. (1996) выделили ген CYCLOIDEA (CYC) , который экспрессируется только в спинных частях цветка львиный зев и отвечает за регуляцию зигоморфных цветков.![]() Двойной мутант CYC и его близкий гомолог DICHOTOMA приводит к радиально-симметричным цветкам львиного зева (Luo et al., 1996). Cubas et al. (1999b) обнаружили, что гомолог гена CYC также отвечает за симметрию цветков льняной жабы обыкновенной.Здесь они смогли показать, что ген CYC у пелорических мутантов был экстенсивно метилирован и заглушен (Cubas et al., 1999b). Примерно в то же время Doebley et al. (1995) проанализировали два локуса количественных признаков, которые контролируют морфологические различия между одомашненной кукурузой ( Zea mays ) и ее диким предшественником teosinte. Они обнаружили разветвленную мутацию teosinte 1 ( tb1 ), которая приводит к усилению разрастания боковых побегов, и показали, что разница между кукурузой и вариантом teosinte TB1 заключается в основном в регуляторных областях гена, i .е., тогда как функция остается той же самой, паттерн экспрессии различается у домашней кукурузы и теозинте (Wang et al.
Двойной мутант CYC и его близкий гомолог DICHOTOMA приводит к радиально-симметричным цветкам львиного зева (Luo et al., 1996). Cubas et al. (1999b) обнаружили, что гомолог гена CYC также отвечает за симметрию цветков льняной жабы обыкновенной.Здесь они смогли показать, что ген CYC у пелорических мутантов был экстенсивно метилирован и заглушен (Cubas et al., 1999b). Примерно в то же время Doebley et al. (1995) проанализировали два локуса количественных признаков, которые контролируют морфологические различия между одомашненной кукурузой ( Zea mays ) и ее диким предшественником teosinte. Они обнаружили разветвленную мутацию teosinte 1 ( tb1 ), которая приводит к усилению разрастания боковых побегов, и показали, что разница между кукурузой и вариантом teosinte TB1 заключается в основном в регуляторных областях гена, i .е., тогда как функция остается той же самой, паттерн экспрессии различается у домашней кукурузы и теозинте (Wang et al. , 1999).
, 1999).
Kosugi et al. (1995) обнаружили, что два промоторных мотива, которые важны для регуляции транскрипции гена ядерного антигена пролиферирующих клеток (PCNA) у риса ( Oryza sativa ), связаны с двумя факторами транскрипции, которые были обозначены как PCF1 и PCF2 (Kosugi и Ohashi, 1997). Наконец, Cubas et al. (1999a) определили, что описанные выше белки T, B1, C, YC и P, CF1 и PCF2 имеют общий консервативный неканонический участок bHLH, одноименный домен TCP (Kosugi and Ohashi, 1997).
Форма и функция факторов транскрипции TCP
Принимая во внимание, что этот обзор будет в основном сосредоточен на эволюционно сохраняющейся роли TCPs в регуляции развития растений и их взаимодействии с эндогенными сигналами и сигналами окружающей среды, крайне важно понять, как они функционируют. Факторы транскрипции TCP делятся на два класса: TCP класса I и класса II. Эти классы различаются составом соответствующих NLS, длиной второй спирали в домене bHLH и наличием богатого аргинином домена с неизвестной функциональностью за пределами домена bHLH (Cubas et al. , 1999а). Этот так называемый R-домен не обнаруживается в TCP класса I, и было предсказано, что он образует гидрофильную α-спираль или структуру спиральной спирали, которая опосредует межбелковые взаимодействия (Lupas et al., 1991; Cubas et al., 1999a). .
Основная область домена TCP важна для связывания ДНК. Замена консервативной пары глицин-пролин в основной области двумя лизинами полностью устраняет ДНК-связывающую активность TCP4 в исследованиях сдвига электрофоретической подвижности (Aggarwal et al., 2010).Добавление связывающего большую бороздку красителя метилового зеленого снижает связывание TCP4 с ДНК, указывая тем самым, что TCP4 связывается с большой бороздкой в двухцепочечной ДНК (Aggarwal et al., 2010).
В различных экспериментальных подходах было показано, что TCP-белки класса I и класса II распознают GC-богатые последовательности в промоторах целевых генов (Kosugi and Ohashi, 1997; Li et al., 2005; Viola et al., 2011; Danisman et al. ., 2012). Различия между предпочтениями связывания класса I и класса II зависят от присутствия глицина или аспарагиновой кислоты в положениях 11 или 15 соответственно (Viola et al. , 2012). Интересно, что последовательности консенсусных сайтов связывания классов I и II не исключают друг друга, что указывает на то, что по крайней мере подмножество потенциальных генов-мишеней нацелено на TCP-белки как класса I, так и класса II. Это привело к предположениям о возможной антагонистической связи между TCP класса I и класса II, где эти белки конкурируют за общие гены-мишени и ингибируют или активируют экспрессию гена в зависимости от того, какой класс доминирует в промоторе гена-мишени (Li et al., 2005). До сих пор это было показано только в одном случае, когда фактор транскрипции TCP Arabidopsis класса I TCP20 связывается с тем же промотором, что и TCP4 класса II, и регулирует целевой ген LIPOXYGENASE2 (LOX2) в направлении, противоположном TCP4 ( Данисман и др., 2012). Однако вероятно, что в будущем будет обнаружено больше случаев антагонизма TCP класса I-класса II, поскольку часто обнаруживается, что эти два класса участвуют в одних и тех же биологических процессах.
, 2012). Интересно, что последовательности консенсусных сайтов связывания классов I и II не исключают друг друга, что указывает на то, что по крайней мере подмножество потенциальных генов-мишеней нацелено на TCP-белки как класса I, так и класса II. Это привело к предположениям о возможной антагонистической связи между TCP класса I и класса II, где эти белки конкурируют за общие гены-мишени и ингибируют или активируют экспрессию гена в зависимости от того, какой класс доминирует в промоторе гена-мишени (Li et al., 2005). До сих пор это было показано только в одном случае, когда фактор транскрипции TCP Arabidopsis класса I TCP20 связывается с тем же промотором, что и TCP4 класса II, и регулирует целевой ген LIPOXYGENASE2 (LOX2) в направлении, противоположном TCP4 ( Данисман и др., 2012). Однако вероятно, что в будущем будет обнаружено больше случаев антагонизма TCP класса I-класса II, поскольку часто обнаруживается, что эти два класса участвуют в одних и тех же биологических процессах.
Подобно многим семействам факторов транскрипции, TCP требует димеризации для связывания с ДНК, поскольку добавление дезоксихолата, ингибитора белок-белковых взаимодействий, к анализам сдвига электрофоретической подвижности приводит к снижению связывания TCP с последовательностями-мишенями (Trémousaygue et al., 2003). Димеризация между факторами транскрипции TCP впервые была описана между PCF1 и PCF2 у риса, которые образуют гомо- и гетеродимеры (Kosugi and Ohashi, 1997). В то время как гомодимер TCP20, например, не связывается с промотором регулятора гомеостаза железа BHLh49 в экспериментах с одним гибридом дрожжей, гетеродимер TCP20 с TCP8 или TCP21 связывает (Andriankaja et al., 2014). Систематический двухгибридный подход дрожжей между TCP Arabidopsis показал, что между TCP возможны многие белок-белковые взаимодействия и что предпочтение отдается связыванию с TCP собственного класса, т.е.е. TCP класса I предпочтительно взаимодействуют с TCP класса I, а TCP класса II предпочтительно взаимодействуют с TCP класса II (Danisman et al. , 2013). Димеризации TCPs способствует IDR (Valsecchi et al., 2013). Они характеризуются низкой компактностью, низкой глобулярностью и более высокой структурной гибкостью и обычно широко присутствуют в факторах транскрипции эукариот (Liu et al., 2006). C-терминальный IDR TCP8 необходим для самосборки TCP8 в димеры и комплексы более высокого порядка. Эти IDR потенциально способствуют гибкости TCP в выборе взаимодействующих партнеров и, таким образом, увеличивают количество потенциальных функций, в которых могут быть задействованы факторы транскрипции TCP, по мнению Thieulin-Pardo et al.(2015). TCP не только взаимодействуют с TCP: были описаны белок-белковые взаимодействия с множеством других белков, включая негативные регуляторы эффекторного иммунитета (Kim et al., 2014), компоненты циркадных часов (Pruneda-Paz et al. , 2009, 2014; Giraud et al., 2010) и др. (Trémousaygue et al., 2003; Tao et al., 2013).
, 2013). Димеризации TCPs способствует IDR (Valsecchi et al., 2013). Они характеризуются низкой компактностью, низкой глобулярностью и более высокой структурной гибкостью и обычно широко присутствуют в факторах транскрипции эукариот (Liu et al., 2006). C-терминальный IDR TCP8 необходим для самосборки TCP8 в димеры и комплексы более высокого порядка. Эти IDR потенциально способствуют гибкости TCP в выборе взаимодействующих партнеров и, таким образом, увеличивают количество потенциальных функций, в которых могут быть задействованы факторы транскрипции TCP, по мнению Thieulin-Pardo et al.(2015). TCP не только взаимодействуют с TCP: были описаны белок-белковые взаимодействия с множеством других белков, включая негативные регуляторы эффекторного иммунитета (Kim et al., 2014), компоненты циркадных часов (Pruneda-Paz et al. , 2009, 2014; Giraud et al., 2010) и др. (Trémousaygue et al., 2003; Tao et al., 2013).
Эволюционно сохраненные роли TCP
Три одноименных белка TCP были охарактеризованы как регуляторы ветвления, симметрии цветков и клеточного цикла (Doebley et al. , 1995; Луо и др., 1996). Позже было показано, что как CYC-подобные, так и PCF-подобные TCPs участвуют в развитии листьев (Kosugi and Ohashi, 1997; Palatnik et al., 2003). С тех пор исследования TCP были сосредоточены на этих трех процессах развития, в основном на выявлении эволюционно консервативных процессов в широком спектре видов растений и на роли регуляции клеточного цикла в наблюдаемых фенотипах. Однако недавно стало ясно, что TCPs не ограничиваются ветвлением, симметрией цветков и развитием листьев, и они также не ограничиваются регуляцией роста, опосредованной клеточным циклом.Оба будут рассмотрены ниже.
, 1995; Луо и др., 1996). Позже было показано, что как CYC-подобные, так и PCF-подобные TCPs участвуют в развитии листьев (Kosugi and Ohashi, 1997; Palatnik et al., 2003). С тех пор исследования TCP были сосредоточены на этих трех процессах развития, в основном на выявлении эволюционно консервативных процессов в широком спектре видов растений и на роли регуляции клеточного цикла в наблюдаемых фенотипах. Однако недавно стало ясно, что TCPs не ограничиваются ветвлением, симметрией цветков и развитием листьев, и они также не ограничиваются регуляцией роста, опосредованной клеточным циклом.Оба будут рассмотрены ниже.
TEOSINTE BRANCHED 1, CYCLOIDEA, факторы транскрипции PCF1 принадлежат к эволюционно консервативному семейству, которое впервые появляется у пресноводных водорослей семейства Charophyta (Navaud et al., 2007). У мохообразных Physcomitrella patens нокаут фактора транскрипции TCP Pp TCP5 приводит к увеличению числа спорангиев, прикрепленных к одной щетинке, что напоминает фенотипов ветвления мутантов tcp у высших наземных растений (Ortiz- Рамирес и др. , 2016). Следовательно, контроль меристематической активности аксиллярных меристем с последующим влиянием на паттерны ветвления, по-видимому, является древней ролью факторов транскрипции TCP (Ortiz-Ramírez et al., 2016). В соответствии с этим открытием фенотипы ветвления проявляются как у однодольных, так и у двудольных растений. Сверхэкспрессия OsTB1 риса, ортолога TB1 кукурузы, привела к сильному снижению количества побегов. На количество пазушных почек у этих растений не повлияло, но их рост был Takeda et al.(2003). Это согласуется с наблюдением, что TCPs влияет не на формирование подмышечных меристем, а на их рост (Braun et al., 2012). Это было показано на горохе (Braun et al., 2012), тополе (Muhr et al., 2016), Arabidopsis (Aguilar-Martínez et al., 2007; Poza-Carrión et al., 2007) и картофеле ( Николас и др., 2015).
, 2016). Следовательно, контроль меристематической активности аксиллярных меристем с последующим влиянием на паттерны ветвления, по-видимому, является древней ролью факторов транскрипции TCP (Ortiz-Ramírez et al., 2016). В соответствии с этим открытием фенотипы ветвления проявляются как у однодольных, так и у двудольных растений. Сверхэкспрессия OsTB1 риса, ортолога TB1 кукурузы, привела к сильному снижению количества побегов. На количество пазушных почек у этих растений не повлияло, но их рост был Takeda et al.(2003). Это согласуется с наблюдением, что TCPs влияет не на формирование подмышечных меристем, а на их рост (Braun et al., 2012). Это было показано на горохе (Braun et al., 2012), тополе (Muhr et al., 2016), Arabidopsis (Aguilar-Martínez et al., 2007; Poza-Carrión et al., 2007) и картофеле ( Николас и др., 2015).
TCP на развитие цветков был продемонстрирован на широком спектре видов растений, включая Arabidopsis , Antirrhinum, однолетний Candytuft ( Iberis amara ) (Busch and Zachgo, 2007; Busch et al. , 2012), покрытосеменных, таких как Aristolochia arborea и Saruma henryi (Horn et al., 2015), виды герберы (Broholm et al., 2008), рис (Yuan et al., 2009), подсолнухи (Fambrini et al. ., 2012), горох (Wang et al., 2008), полынь (Kim et al., 2008), жимолость Морроу ( Lonicera morrowii ) (Howarth, Donoghue, 2006), Knautia macedonica (Berger et al. , 2016) и орхидеи (De Paolo et al., 2015).
, 2012), покрытосеменных, таких как Aristolochia arborea и Saruma henryi (Horn et al., 2015), виды герберы (Broholm et al., 2008), рис (Yuan et al., 2009), подсолнухи (Fambrini et al. ., 2012), горох (Wang et al., 2008), полынь (Kim et al., 2008), жимолость Морроу ( Lonicera morrowii ) (Howarth, Donoghue, 2006), Knautia macedonica (Berger et al. , 2016) и орхидеи (De Paolo et al., 2015).
Филогенетический анализ показал, что CYCLOIDEA-подобные TCPs претерпели два основных события дупликации, которые оба предшествовали образованию основных эвдикотов (Howarth and Donoghue, 2006).В Arabidopsis все три клада CYC представлены TCP12, TCP1 и TCP18 соответственно (Howarth and Donoghue, 2006). В частности, кладка CYC2, представленная TCP1 в Arabidopsis , претерпела несколько дополнительных дупликаций и была изучена на предмет ее влияния на симметрию цветков, поскольку она содержит исходный ген CYC Antirrhinum (Howarth and Donoghue, 2006). Интересное замечание заключается в том, что дупликация TCP, подобных CYCLOIDEA, почти совпадает с основными событиями дупликации гомеотических факторов транскрипции MADS-бокса APETALA3, AGAMOUS и SEPALLATA, все три из которых являются важными факторами для определения идентичности органов у цветковых растений. (Ховарт и Донохью, 2006).Это говорит о том, что генетические компоненты, которые важны для определения органов цветков, диверсифицировались в то же время, что и компоненты, важные для регуляции их роста. Факторы транскрипции TCP были идентифицированы как мишени для Arabidopsis APETALA1 и SEPALLATA3 (Kaufmann et al., 2009, 2010), что подчеркивает возможную связь между формированием идентичности органов и регуляцией роста между факторами транскрипции MADS-бокса и TCP (Dornelas et al. , 2011).
Интересное замечание заключается в том, что дупликация TCP, подобных CYCLOIDEA, почти совпадает с основными событиями дупликации гомеотических факторов транскрипции MADS-бокса APETALA3, AGAMOUS и SEPALLATA, все три из которых являются важными факторами для определения идентичности органов у цветковых растений. (Ховарт и Донохью, 2006).Это говорит о том, что генетические компоненты, которые важны для определения органов цветков, диверсифицировались в то же время, что и компоненты, важные для регуляции их роста. Факторы транскрипции TCP были идентифицированы как мишени для Arabidopsis APETALA1 и SEPALLATA3 (Kaufmann et al., 2009, 2010), что подчеркивает возможную связь между формированием идентичности органов и регуляцией роста между факторами транскрипции MADS-бокса и TCP (Dornelas et al. , 2011).
В Antirrhinum CYC регулирует симметрию через фактор транскрипции Myb-домена RADIALIS (Corley et al., 2005). Избыточная экспрессия CYC в Arabidopsis приводит к более крупным лепесткам, содержащим увеличенные лепестковые клетки (Costa et al. , 2005). Регулирование роста цветков не ограничивается CYC-подобными TCP класса II. У мутанта jaw -D развитие лепестков отличается от Arabidopsis дикого типа (Palatnik et al., 2003). Nag et al. (2009) показали, что это зависит от регуляции miR319 TCP4 . Устойчивая к микроРНК форма TCP4 под контролем промотора APETALA3 экспрессируется только в органах цветков и приводит к значительно меньшим цветам, которые состоят только из плодолистиков и чашелистиков, без каких-либо лепестков или тычинок, тогда как проростки этих растений выглядят нормально (Nag et al., 2009).
, 2005). Регулирование роста цветков не ограничивается CYC-подобными TCP класса II. У мутанта jaw -D развитие лепестков отличается от Arabidopsis дикого типа (Palatnik et al., 2003). Nag et al. (2009) показали, что это зависит от регуляции miR319 TCP4 . Устойчивая к микроРНК форма TCP4 под контролем промотора APETALA3 экспрессируется только в органах цветков и приводит к значительно меньшим цветам, которые состоят только из плодолистиков и чашелистиков, без каких-либо лепестков или тычинок, тогда как проростки этих растений выглядят нормально (Nag et al., 2009).
Репрессор транскрипции «цинковые пальцы» RABBIT EARS контролирует экспрессию TCP TCP5, TCP13 и TCP17 и неправильную экспрессию как RABBIT EARS , так и этих TCP , что приводит к аномальному развитию лепестков у Arabidopsis и Arabidopsis. Ирландский, 2015). Репрессия этих TCP ведет к ранней остановке митотической активности во время развития лепестков (Huang and Irish, 2015). Интересно, что обратное происходит при подавлении TCP5, TCP13 и TCP17 в листьях, где клетки листа продолжают митотические деления дольше, чем у растений дикого типа (Efroni et al., 2008). Здесь влияние факторов транскрипции TCP на развитие органов зависит от контекста органа. Это подчеркивает важность регуляторного взаимодействия между TCP и регуляторами идентичности органов. Хотя есть намеки на это взаимодействие между TCPs и факторами транскрипции MADS box во время развития цветков, такое взаимодействие еще предстоит показать во время развития др. Органов (Dornelas et al., 2011).
Интересно, что обратное происходит при подавлении TCP5, TCP13 и TCP17 в листьях, где клетки листа продолжают митотические деления дольше, чем у растений дикого типа (Efroni et al., 2008). Здесь влияние факторов транскрипции TCP на развитие органов зависит от контекста органа. Это подчеркивает важность регуляторного взаимодействия между TCP и регуляторами идентичности органов. Хотя есть намеки на это взаимодействие между TCPs и факторами транскрипции MADS box во время развития цветков, такое взаимодействие еще предстоит показать во время развития др. Органов (Dornelas et al., 2011).
Первые указания на роль TCP в развитии листа получены из работы Antirrhinum (Nath et al., 2003). Мутант TCP Antirrhinum class II cin демонстрирует морщинистые листья, которые являются результатом изменения регуляции клеточного цикла во время развития листа (Nath et al., 2003). По сути, митотические деления развивающихся листовых клеток на кончике листа задерживаются первыми, а те, что у основания листа, задерживаются в последнюю очередь. Результатом этого последовательного действия по задержке является так называемый фронт остановки, который перемещается от кончика листа к основанию. Форма этого фронта остановки отличается в cin листьях, чем в листьях дикого типа, что приводит к измененной кривизне листа (Nath et al., 2003). У Arabidopsis подобное поведение наблюдается у мутанта jaw -D (Palatnik et al., 2003). Jaw -D является сверхэкспрессором микроРНК miR319a, в которой CIN -подобные TCP класса II TCP2, TCP3, TCP4, TCP10 и TCP24 подавлены (Palatnik et al., 2003). Jaw Мутанты -D обнаруживают зазубренные листья, аномальные лепестки и замедленное развитие и старение листьев (Palatnik et al., 2003). Этот фенотип обусловлен задержкой развития листьев, при которой фронт митотической остановки начинается позже, чем у растений дикого типа (Efroni et al., 2008). Недавно было показано, что регулируемые miR319a факторы транскрипции TCP действуют избыточно с факторами транскрипции NGATHA, чтобы ограничивать меристематическую активность меристем листа во время развития листа (Alvarez et al.
Результатом этого последовательного действия по задержке является так называемый фронт остановки, который перемещается от кончика листа к основанию. Форма этого фронта остановки отличается в cin листьях, чем в листьях дикого типа, что приводит к измененной кривизне листа (Nath et al., 2003). У Arabidopsis подобное поведение наблюдается у мутанта jaw -D (Palatnik et al., 2003). Jaw -D является сверхэкспрессором микроРНК miR319a, в которой CIN -подобные TCP класса II TCP2, TCP3, TCP4, TCP10 и TCP24 подавлены (Palatnik et al., 2003). Jaw Мутанты -D обнаруживают зазубренные листья, аномальные лепестки и замедленное развитие и старение листьев (Palatnik et al., 2003). Этот фенотип обусловлен задержкой развития листьев, при которой фронт митотической остановки начинается позже, чем у растений дикого типа (Efroni et al., 2008). Недавно было показано, что регулируемые miR319a факторы транскрипции TCP действуют избыточно с факторами транскрипции NGATHA, чтобы ограничивать меристематическую активность меристем листа во время развития листа (Alvarez et al. , 2016). Этот фенотип также был очевиден у растений, экспрессирующих искусственную микроРНК против TCP класса II TCP5, TCP13 и TCP17 , и фенотип был чрезвычайно сильным, когда эти растения были скрещены с растениями jaw -D (Efroni et al., 2008 г.).
, 2016). Этот фенотип также был очевиден у растений, экспрессирующих искусственную микроРНК против TCP класса II TCP5, TCP13 и TCP17 , и фенотип был чрезвычайно сильным, когда эти растения были скрещены с растениями jaw -D (Efroni et al., 2008 г.).
класса II также регулируют развитие листьев в сложных листьях томатов. Ортологом Arabidopsis miR319-чувствительных TCPs у томатов является LA , и он находится под контролем miR319 томата (Ori et al., 2007). Мутанты La демонстрируют простые листья, тогда как сверхэкспрессия miR319 без нечувствительности LA к микроРНК приводит к усиленному разделению сложных листьев. Кроме того, листья томатов со сверхэкспрессией miR319 вырастают на 3 месяца дольше, чем листья дикого типа, и демонстрируют признаки поздней дифференцировки, что является поведением, идентичным поведению растений Arabidopsis jaw -D (Ori et al., 2007; Efroni et al., 2008). Избыточная экспрессия miR319 в однодольных Agrostis stolonifera (ползучая полевица) ведет к подавлению регуляции TCP класса II и к образованию более широких и толстых листьев, которые отличаются от листьев дикого типа (Zhou et al. , 2013). Этот фенотип проистекает из увеличенного количества клеток в трансгенной полевице, подобно jaw -D у Arabidopsis (Efroni et al., 2008; Zhou et al., 2013). В целом экспрессия CIN-подобных генов тесно коррелирует с формой листьев как у видов Solanaceae, так и у тополя пустынного ( Populus euphratica ) (Shleizer-Burko et al., 2011; Ma et al., 2016).
, 2013). Этот фенотип проистекает из увеличенного количества клеток в трансгенной полевице, подобно jaw -D у Arabidopsis (Efroni et al., 2008; Zhou et al., 2013). В целом экспрессия CIN-подобных генов тесно коррелирует с формой листьев как у видов Solanaceae, так и у тополя пустынного ( Populus euphratica ) (Shleizer-Burko et al., 2011; Ma et al., 2016).
Экспрессия TCP3 с доминантным репрессорным доменом привела к серьезным нарушениям развития Arabidopsis во всех органах (Koyama et al., 2007), включая образование эктопических побегов, зубчатых листьев, модифицированных чашелистиков и лепестков и образование волнистой кремнистой жидкости. Это было связано с неправильной экспрессией граничных специфических генов, то есть CUC и LATERAL ORGAN BOUNDARIES (Koyama et al., 2007). Также в Antirrhinum было обнаружено, что ортолог Arabidopsis TCP15 взаимодействует с CUPULIFORMIS, белком, который связан с белками CUC Arabidopsis (Weir et al. , 2004). Кроме того, два TCP14 и TCP15 Arabidopsis класса I оказались избыточными в влиянии на пролиферацию клеток во время развития листа и в других тканях у Arabidopsis . Однако наиболее очевидный эффект был замечен в длине междоузлий, которая уменьшается у мутантов tcp14 tcp15 и приводит к более коротким растениям (Kieffer et al., 2011).
, 2004). Кроме того, два TCP14 и TCP15 Arabidopsis класса I оказались избыточными в влиянии на пролиферацию клеток во время развития листа и в других тканях у Arabidopsis . Однако наиболее очевидный эффект был замечен в длине междоузлий, которая уменьшается у мутантов tcp14 tcp15 и приводит к более коротким растениям (Kieffer et al., 2011).
В то время как функции TCP, таким образом, были очень хорошо охарактеризованы в этих ветвлениях, развитии цветков и листьев для широкого спектра видов растений (рис. 1), есть намеки на то, что это всего лишь подмножество ролей TCP в развитии.Было показано, что TCPs активируются при впитывании сухих семян, и прорастание линий вставки транспозона tcp14 , по-видимому, было ниже, чем в семенах дикого типа (Tatematsu et al., 2008). Хотя здесь экспрессия TCP14 в линиях транспозонов не обязательно была ниже, чем у дикого типа, что указывает на то, что TCP14 не может быть единственной причиной снижения скорости прорастания (Tatematsu et al. , 2008). Подавление экспрессии TCP в хлопке привело к уменьшению длины волокон хлопка, а также к увеличению количества боковых отростков и задержке роста, что свидетельствует о снижении апикального доминирования (Hao et al., 2012). Сверхэкспрессия miR319 в китайской капусте не только приводила к измененному развитию листьев, но и кочаны капусты были более округлыми, чем у капусты с низкой экспрессией miR319 и более высокой экспрессией ее целевого гена BrpTCP4-1 (Mao et al., 2014). Гетерологичная экспрессия OsTCP19 риса в Arabidopsis привела к меньшему количеству боковых корней (Mukhopadhyay et al., 2015). В огурце мутация гена TCP привела к уникальному фенотипу растения. У пораженных растений огурца не появились усики, а появились побеги.Авторы этого исследования предполагают, что здесь TCPs не только влияют на рост органа, но также определяют идентичность органа (Wang S. et al., 2015). Похожий фенотип был обнаружен у дынь, где однонуклеотидная мутация в CmTCP1 привела к мутации без усиков Chiba.
, 2008). Подавление экспрессии TCP в хлопке привело к уменьшению длины волокон хлопка, а также к увеличению количества боковых отростков и задержке роста, что свидетельствует о снижении апикального доминирования (Hao et al., 2012). Сверхэкспрессия miR319 в китайской капусте не только приводила к измененному развитию листьев, но и кочаны капусты были более округлыми, чем у капусты с низкой экспрессией miR319 и более высокой экспрессией ее целевого гена BrpTCP4-1 (Mao et al., 2014). Гетерологичная экспрессия OsTCP19 риса в Arabidopsis привела к меньшему количеству боковых корней (Mukhopadhyay et al., 2015). В огурце мутация гена TCP привела к уникальному фенотипу растения. У пораженных растений огурца не появились усики, а появились побеги.Авторы этого исследования предполагают, что здесь TCPs не только влияют на рост органа, но также определяют идентичность органа (Wang S. et al., 2015). Похожий фенотип был обнаружен у дынь, где однонуклеотидная мутация в CmTCP1 привела к мутации без усиков Chiba. Также здесь усики были преобразованы в побеговые и листоподобные структуры (Mizuno et al., 2015). Это было бы первым признаком того, что TCP могут действовать как регуляторы идентичности органов. Дальнейшие исследования еще не раскрыли, является ли функция TCP в регуляции идентичности органов усиков уникальной и новой ролью или другим органам растений также необходимы TCP для определения своей идентичности.
Также здесь усики были преобразованы в побеговые и листоподобные структуры (Mizuno et al., 2015). Это было бы первым признаком того, что TCP могут действовать как регуляторы идентичности органов. Дальнейшие исследования еще не раскрыли, является ли функция TCP в регуляции идентичности органов усиков уникальной и новой ролью или другим органам растений также необходимы TCP для определения своей идентичности.
РИСУНОК 1. Филогенетическое дерево видов растений, в ветвлении которых задействованы факторы транскрипции TCP (Takeda et al., 2003; Aguilar-Martínez et al., 2007; Poza-Carrión et al., 2007; Bai et al. ., 2012; Braun et al., 2012; Drummond et al., 2015; Nicolas et al., 2015; Muhr et al., 2016) (синие точки), развитие цветков (Linnaeus, Rudberg, 1744; Keeble et al. , 1910; Corley et al., 2005; Costa et al., 2005; Busch, Zachgo, 2007; Broholm et al., 2008; Kim et al., 2008; Наг и др., 2009; Юань и др., 2009; Ховарт и др., 2011; Busch et al., 2012; Тяхтихарью и др. , 2012; Класен-Бокхофф и др., 2013; Juntheikki-Palovaara et al., 2014; Де Паоло и др., 2015; Хорн и др., 2015; Lucero et al., 2015; Ван и др., 2008; Ван X. и др., 2015; Ян и др., 2015; Berger et al., 2016) (фиолетовые точки) или развитие листьев (Kosugi, Ohashi, 1997; Nath et al., 2003; Palatnik et al., 2003; Koyama et al., 2007, 2010a, b; Ori et al. , 2007; Efroni et al., 2008; Kieffer et al., 2011; Mimida et al., 2011; Сарвепалли и Нат, 2011; Данисман и др., 2012, 2013; Агилар-Мартинес и Синха, 2013 г .; Бурко и др., 2013; Тао и др., 2013; Чжоу и др., 2013; Баллестер и др., 2015; Хуанг и Ирландский, 2015; Ma et al., 2016) (зеленые точки) соответственно. Филогенетическое дерево было создано с использованием Phylotree и iTOL (Letunic and Bork, 2016).
, 2012; Класен-Бокхофф и др., 2013; Juntheikki-Palovaara et al., 2014; Де Паоло и др., 2015; Хорн и др., 2015; Lucero et al., 2015; Ван и др., 2008; Ван X. и др., 2015; Ян и др., 2015; Berger et al., 2016) (фиолетовые точки) или развитие листьев (Kosugi, Ohashi, 1997; Nath et al., 2003; Palatnik et al., 2003; Koyama et al., 2007, 2010a, b; Ori et al. , 2007; Efroni et al., 2008; Kieffer et al., 2011; Mimida et al., 2011; Сарвепалли и Нат, 2011; Данисман и др., 2012, 2013; Агилар-Мартинес и Синха, 2013 г .; Бурко и др., 2013; Тао и др., 2013; Чжоу и др., 2013; Баллестер и др., 2015; Хуанг и Ирландский, 2015; Ma et al., 2016) (зеленые точки) соответственно. Филогенетическое дерево было создано с использованием Phylotree и iTOL (Letunic and Bork, 2016).
Влияние функций TCP на цикл ячейки — прямое или косвенное?
Ранее в исследовании TCP основное внимание уделялось клеточному циклу как основной мишени регуляции TCP (Kosugi and Ohashi, 1997; Li et al., 2005). В то время как связывание с генами клеточного цикла было показано в определенных случаях (Li et al.![]() , 2005; Davière et al., 2014), тщательный анализ паттернов клеточного деления и изменений транскриптов во время развития jaw -D листа показал, что класс II-TCP-зависимая регуляция клеточного цикла может быть косвенной (Efroni et al., 2008). Кроме того, связывание TCP TCP20 класса I с генами клеточного цикла было показано только один раз и in vitro (Li et al., 2005), тогда как прямой анализ целевого гена показывает, что синтез гормона, особенно синтез жасмоната, прямо нацелен на TCP20 (Данисман и др., 2012). И TCP4, и TCP20 влияют на развитие листьев посредством синтеза метилжасмоната, гормона, который выполняет множество функций в развитии растений и ответе на раны и патогены (Schommer et al., 2008). Жасмонат, обычно известный своей ролью в ранении и ответной реакции патогенов, также влияет на клеточный цикл (Świątek et al., 2002).
, 2005; Davière et al., 2014), тщательный анализ паттернов клеточного деления и изменений транскриптов во время развития jaw -D листа показал, что класс II-TCP-зависимая регуляция клеточного цикла может быть косвенной (Efroni et al., 2008). Кроме того, связывание TCP TCP20 класса I с генами клеточного цикла было показано только один раз и in vitro (Li et al., 2005), тогда как прямой анализ целевого гена показывает, что синтез гормона, особенно синтез жасмоната, прямо нацелен на TCP20 (Данисман и др., 2012). И TCP4, и TCP20 влияют на развитие листьев посредством синтеза метилжасмоната, гормона, который выполняет множество функций в развитии растений и ответе на раны и патогены (Schommer et al., 2008). Жасмонат, обычно известный своей ролью в ранении и ответной реакции патогенов, также влияет на клеточный цикл (Świątek et al., 2002).
Жасмонат — не единственный гормон растения, который может опосредовать регуляцию TCP клеточного цикла. Доказательства участия гормонов в TCP-опосредованной регуляции роста накопились в последние годы (Nicolas and Cubas, 2016). Функции TCP были связаны с абсцизовой кислотой (Tatematsu et al., 2008; González-Grandío et al., 2013; Mukhopadhyay et al., 2015), ауксином (Kosugi et al., 1995; Ben-Gera and Ori, 2012; Uberti-Manassero et al., 2012; Das Gupta et al., 2014), брассиностероид (Guo et al., 2010), цитокинин (Steiner et al., 2012; Efroni et al., 2013), GA (Yanai et al. ., 2011; Das Gupta et al., 2014; Davière et al., 2014), жасмоновая кислота (Schommer et al., 2008; Danisman et al., 2012), салициловая кислота (Wang X. et al., 2015) и сигнальные пути стриголактона (Dun et al., 2012; Hu et al., 2014) (рисунок 2).
Функции TCP были связаны с абсцизовой кислотой (Tatematsu et al., 2008; González-Grandío et al., 2013; Mukhopadhyay et al., 2015), ауксином (Kosugi et al., 1995; Ben-Gera and Ori, 2012; Uberti-Manassero et al., 2012; Das Gupta et al., 2014), брассиностероид (Guo et al., 2010), цитокинин (Steiner et al., 2012; Efroni et al., 2013), GA (Yanai et al. ., 2011; Das Gupta et al., 2014; Davière et al., 2014), жасмоновая кислота (Schommer et al., 2008; Danisman et al., 2012), салициловая кислота (Wang X. et al., 2015) и сигнальные пути стриголактона (Dun et al., 2012; Hu et al., 2014) (рисунок 2).
РИСУНОК 2. Гормональные пути, связанные с Arabidopsis факторов транскрипции TCP и ортологов. Белки были построены в соответствии с их филогенетическим сходством с использованием PhyML и TreeDyne (Dereeper et al., 2008). АК, абсцизовая кислота; AX, ауксин; BR, брассиностероиды; СК, цитокинин; ЕТ, этилен; GA, гибберелловая кислота; JA, жасмоновая кислота; SA, салициловая кислота; SL, стриголактоны.
Помимо гормонального контроля роста, факторы транскрипции TCP также участвуют в других биологических процессах, которые, в свою очередь, влияют на рост.Напр., Сайты связывания факторов транскрипции TCP были идентифицированы в промоторах генов CYTOCHROME C1, и 103, которые кодируют компоненты механизма окислительного фосфорилирования митохондрий и биогенеза белков (Welchen and Gonzalez, 2006). Авторы этого исследования предположили, что факторы транскрипции TCP, связывающие эти сайты, координируют генезис митохондрий и функцию их роста в новых органах (Welchen and Gonzalez, 2006). Другое исследование показало, что эти гены содержат в своих промоторах элемент GGGC (C / T), который важен для суточной регуляции экспрессии их генов (Giraud et al., 2010). Эти промоторы связаны с факторами транскрипции TCP, что указывает на роль в суточной регуляции транскриптов механизма окислительного фосфорилирования митохондрий (Giraud et al., 2010). Ранее было обнаружено, что TCP21 связывается с промотором гена основных часов CCA1 и регулирует его экспрессию (Pruneda-Paz et al. , 2009). TCP21 служит ингибитором CCA1 в течение дня, а димеризация TOC1 с TCP21 отменяет его связывание с промотором CCA1 .В двойном мутанте с часовым геном LHY tcp21 / lhy значительно сокращает период экспрессии CCA1 (Pruneda-Paz et al., 2009). Не только TCP21, но и другие TCP связываются с CCA1 в исследованиях на дрожжах и экспериментах по коиммунопреципитации (Giraud et al., 2010; Pruneda-Paz et al., 2014). Недавнее исследование также показало, что TCP20 и TCP22 действуют как активаторы CCA1 утром, выполняя важную роль в кругообороте циркадных часов (Wu et al., 2016). Это означает, что белки TCP связываются с промоторами часовых генов, регулируют их экспрессию, димеризуются с часовыми белками и связываются с нижележащими целями часов (Pruneda-Paz et al., 2009, 2014; Giraud et al., 2010; Wu et al. al., 2016) (рисунок 3). В целом становится ясно, что TCP влияет не только на рост через клеточный цикл. Вместо этого они участвуют в различных биологических процессах, прямо или косвенно влияющих на рост.
, 2009). TCP21 служит ингибитором CCA1 в течение дня, а димеризация TOC1 с TCP21 отменяет его связывание с промотором CCA1 .В двойном мутанте с часовым геном LHY tcp21 / lhy значительно сокращает период экспрессии CCA1 (Pruneda-Paz et al., 2009). Не только TCP21, но и другие TCP связываются с CCA1 в исследованиях на дрожжах и экспериментах по коиммунопреципитации (Giraud et al., 2010; Pruneda-Paz et al., 2014). Недавнее исследование также показало, что TCP20 и TCP22 действуют как активаторы CCA1 утром, выполняя важную роль в кругообороте циркадных часов (Wu et al., 2016). Это означает, что белки TCP связываются с промоторами часовых генов, регулируют их экспрессию, димеризуются с часовыми белками и связываются с нижележащими целями часов (Pruneda-Paz et al., 2009, 2014; Giraud et al., 2010; Wu et al. al., 2016) (рисунок 3). В целом становится ясно, что TCP влияет не только на рост через клеточный цикл. Вместо этого они участвуют в различных биологических процессах, прямо или косвенно влияющих на рост.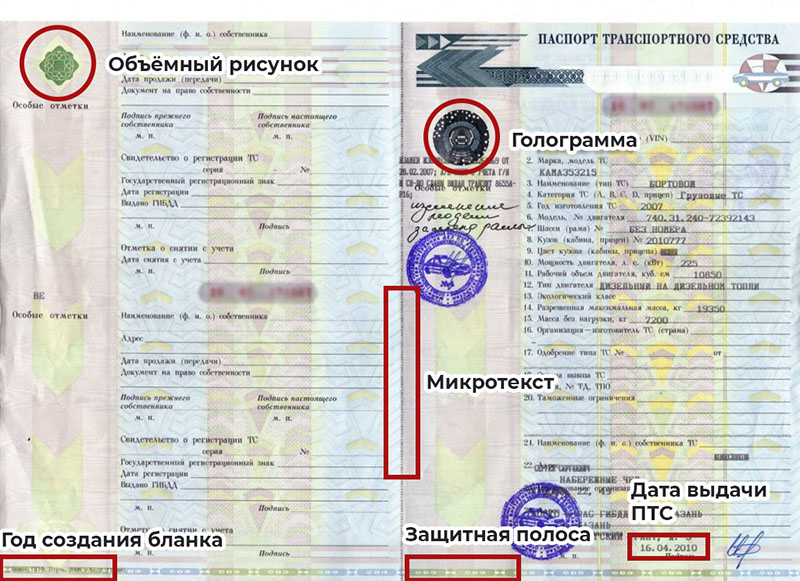
РИСУНОК 3. Взаимодействие факторов транскрипции TCP с компонентами циркадных часов как в схеме центральных часов, так и в последующих процессах.TCP класса I и класса II обозначены зеленым и синим цветом соответственно. Известные компоненты часов показаны серым цветом. Белки представлены кружками, гены — квадратами. Димеры изображены в виде перекрывающихся кругов. CCA1 Ингибирование TCP21 отменяется димеризацией TCP21 с TOC1. Димер CCA1 / LHY ингибирует экспрессию TCP21 (Pruneda-Paz et al., 2009). Эффект девяти TCP, которые связываются с промотором CCA1 , в исследованиях с одним гибридом дрожжей неизвестен (Pruneda-Paz et al., 2014). Ниже часов гетеродимеры TCP / часового компонента регулируют ритмическую экспрессию митохондриальных белков в зависимости от количества и расположения сайтов связывания TCP в промоторах митохондриальных генов (Giraud et al., 2010).
Посредничество сигналов окружающей среды в реакцию роста
Эта картина становится еще более сложной, поскольку TCP также передают сигналы окружающей среды в реакции роста. Было обнаружено, что TCP участвует в защите от патогенов. Во-первых, обширное исследование показало, что инфекция как Pseudomonas syringae , так и Hyaloperonospora arabidopsidis приводит к снижению белка TCP14 (Mukhtar et al., 2011). Секретированные белки патогенных бактерий, перенесенные цикадкой Aster ( Macrosteles quadrilineatus ) на Arabidopsis , были способны димеризоваться и дестабилизировать белки TCP2, TCP4 и TCP7, содержащие оба класса факторов транскрипции TCP (Sugio et al., 2011, 2014). Сверхэкспрессия ответственного фитоплазменного белка SECRETED ASTER YELLOWS-WITCHES BROOM PROTEIN 11 в Arabidopsis дестабилизирует TCP2, TCP3, TCP4, TCP5, TCP10, TCP13, TCP17 и TCP24 и приводит к фенотипам челюсти -D (Sugio ., 2011). Кроме того, уровни жасмоновой кислоты в инфицированных листьях Arabidopsis значительно ниже по сравнению с необработанными листьями, что указывает на то, что защитные механизмы растения снижаются при заражении патогеном.
Было обнаружено, что TCP участвует в защите от патогенов. Во-первых, обширное исследование показало, что инфекция как Pseudomonas syringae , так и Hyaloperonospora arabidopsidis приводит к снижению белка TCP14 (Mukhtar et al., 2011). Секретированные белки патогенных бактерий, перенесенные цикадкой Aster ( Macrosteles quadrilineatus ) на Arabidopsis , были способны димеризоваться и дестабилизировать белки TCP2, TCP4 и TCP7, содержащие оба класса факторов транскрипции TCP (Sugio et al., 2011, 2014). Сверхэкспрессия ответственного фитоплазменного белка SECRETED ASTER YELLOWS-WITCHES BROOM PROTEIN 11 в Arabidopsis дестабилизирует TCP2, TCP3, TCP4, TCP5, TCP10, TCP13, TCP17 и TCP24 и приводит к фенотипам челюсти -D (Sugio ., 2011). Кроме того, уровни жасмоновой кислоты в инфицированных листьях Arabidopsis значительно ниже по сравнению с необработанными листьями, что указывает на то, что защитные механизмы растения снижаются при заражении патогеном. Аналогичный эффект был обнаружен у яблок, где патоген растений Candidatus Phytoplasma mali связывается с двумя факторами транскрипции TCP и вызывает морфогенетические изменения, которые происходят одновременно со снижением уровней жасмоновой кислоты, салициловой кислоты и абсцизовой кислоты (Janik et al., 2016). Дальнейшие исследования идентифицировали TCP8 и TCP9 класса I как важные факторы экспрессии ICS1 , который кодирует ключевой фермент в синтезе салициловой кислоты (Wang X. et al., 2015). В другом исследовании было установлено, что TCP21 связывается с промотором ICS1 , и индукция экспрессии ICS1 салициловой кислотой блокируется у мутантов tcp21 (Zheng et al., 2015). TCP класса I также взаимодействуют с белками, которые, как известно, регулируют экспрессию ICS1 , т.е.е. факторы транскрипции WRKY28, NAC019 и ETHYLENE INSENSITIVE 1 и кальмодулин-связывающий белок. ДЕФИЦИТ СИСТЕМНОЙ ПРИОБРЕТЕННОЙ РЕЗИСТЕНТНОСТИ 1. Следовательно, двойной мутант tcp8 tcp9 демонстрирует повышенную чувствительность к инфекции Pseudomonas syringae pv.
Аналогичный эффект был обнаружен у яблок, где патоген растений Candidatus Phytoplasma mali связывается с двумя факторами транскрипции TCP и вызывает морфогенетические изменения, которые происходят одновременно со снижением уровней жасмоновой кислоты, салициловой кислоты и абсцизовой кислоты (Janik et al., 2016). Дальнейшие исследования идентифицировали TCP8 и TCP9 класса I как важные факторы экспрессии ICS1 , который кодирует ключевой фермент в синтезе салициловой кислоты (Wang X. et al., 2015). В другом исследовании было установлено, что TCP21 связывается с промотором ICS1 , и индукция экспрессии ICS1 салициловой кислотой блокируется у мутантов tcp21 (Zheng et al., 2015). TCP класса I также взаимодействуют с белками, которые, как известно, регулируют экспрессию ICS1 , т.е.е. факторы транскрипции WRKY28, NAC019 и ETHYLENE INSENSITIVE 1 и кальмодулин-связывающий белок. ДЕФИЦИТ СИСТЕМНОЙ ПРИОБРЕТЕННОЙ РЕЗИСТЕНТНОСТИ 1. Следовательно, двойной мутант tcp8 tcp9 демонстрирует повышенную чувствительность к инфекции Pseudomonas syringae pv. maculicola ES4326 (Wang X. et al., 2015). Факторы транскрипции TCP частично контролируют защиту патогенов посредством второго пути, то есть путем противодействия действию SUPPRESSOR OF rps4-RLD1, белка, который негативно регулирует запускаемый эффектором иммунитет у Arabidopsis (Kim et al., 2014). Отсутствие TCP в тройном мутанте tcp8 tcp14 tcp15 приводит к усилению роста Pseudomonas syringae DC3000 по сравнению с растениями дикого типа (Kim et al., 2014).
maculicola ES4326 (Wang X. et al., 2015). Факторы транскрипции TCP частично контролируют защиту патогенов посредством второго пути, то есть путем противодействия действию SUPPRESSOR OF rps4-RLD1, белка, который негативно регулирует запускаемый эффектором иммунитет у Arabidopsis (Kim et al., 2014). Отсутствие TCP в тройном мутанте tcp8 tcp14 tcp15 приводит к усилению роста Pseudomonas syringae DC3000 по сравнению с растениями дикого типа (Kim et al., 2014).
Недавние исследования показали, что факторы транскрипции TCP регулируют время цветения. Нокаут фактора транскрипции TCP класса I TCP23 привел к более раннему цветению, чем у дикого типа, тогда как линии со сверхэкспрессией TCP23 показали замедленное цветение (Balsemão-Pires et al., 2013). Цветочный переход пазушных меристем у Arabidopsis контролируется взаимодействием между белками времени цветения FT и TWIN SISTER OF FT и BRC1 (Niwa et al., 2013). Межбелковые взаимодействия между этими факторами транскрипции были показаны в дрожжевых двугибридных, бимолекулярных флуоресцентных анализах комплементации и in vitro pull-down анализах (Niwa et al. , 2013). Поскольку мутанты brc1 демонстрируют ускоренное цветение, а ft и сестры-близнецы мутантов ft демонстрируют более медленное цветение пазушных меристем, соответственно, кажется, что существует антагонистическая связь между BRC1 и белками времени цветения (Niwa et al., 2013). Вероятно, что димеризация BRC1 с помощью FT и TWIN SISTER OF FT репрессирует их функцию в подмышечных меристемах (Niwa et al., 2013). Ортологи яблочного FT Md FT1 и 2 также, как было обнаружено, взаимодействуют с факторами транскрипции TCP (Mimida et al., 2011). Сверхэкспрессия miR319 томата привела к цветению с меньшим количеством листьев, чем у томата дикого типа, и было показано, что LA связывается с промоторами ортологов APETALA1 и FRUITFUL томата (Burko et al., 2013).
, 2013). Поскольку мутанты brc1 демонстрируют ускоренное цветение, а ft и сестры-близнецы мутантов ft демонстрируют более медленное цветение пазушных меристем, соответственно, кажется, что существует антагонистическая связь между BRC1 и белками времени цветения (Niwa et al., 2013). Вероятно, что димеризация BRC1 с помощью FT и TWIN SISTER OF FT репрессирует их функцию в подмышечных меристемах (Niwa et al., 2013). Ортологи яблочного FT Md FT1 и 2 также, как было обнаружено, взаимодействуют с факторами транскрипции TCP (Mimida et al., 2011). Сверхэкспрессия miR319 томата привела к цветению с меньшим количеством листьев, чем у томата дикого типа, и было показано, что LA связывается с промоторами ортологов APETALA1 и FRUITFUL томата (Burko et al., 2013).
Восприятие соотношения красного и дальнего красного света (R: FR) сообщает растению о затенении соседней растительностью, а более низкое соотношение R: FR приводит к подавлению разрастания подмышечной меристемы, позволяя растению инвестировать в более длинный гипокотиль и в конечном итоге избегать затенение. В Arabidopsis удлинение гипокотиля регулируется посредством фактора транскрипции bHLH PHYTOCHROME INTERACTING FACTOR 4, который, среди прочего, активирует экспрессию YUCCA8 , способствуя удлинению клеток (Sun et al., 2012). YUCCA2, 5 и 8 также являются прямыми генами-мишенями TCP4. Фактически, индуцированная сверхэкспрессия TCP4 приводит к удлинению гипокотилей, и этот эффект зависит от передачи сигналов как ауксина, так и брассиностероида (Challa et al., 2016). В картофеле регулирование BRC1a зависит от R: FR. BRC1a бывает двух видов: короткая (BRC1a S ) и длинная версия с альтернативным соединением (BRC1a L ). Оба приводят к образованию белков, но более короткая форма является цитоплазматической и не связывается с генами-мишенями в ядре.Соотношение между этими двумя формами меняется при обезглавливании побегов картофеля, нахождении в темноте и в условиях низкого R: FR (Nicolas et al., 2015). В то время как декапитация приводит к относительному увеличению BRC1a S , темнота и обработка с низким R: FR приводят к относительному увеличению содержания BRC1a L .
В Arabidopsis удлинение гипокотиля регулируется посредством фактора транскрипции bHLH PHYTOCHROME INTERACTING FACTOR 4, который, среди прочего, активирует экспрессию YUCCA8 , способствуя удлинению клеток (Sun et al., 2012). YUCCA2, 5 и 8 также являются прямыми генами-мишенями TCP4. Фактически, индуцированная сверхэкспрессия TCP4 приводит к удлинению гипокотилей, и этот эффект зависит от передачи сигналов как ауксина, так и брассиностероида (Challa et al., 2016). В картофеле регулирование BRC1a зависит от R: FR. BRC1a бывает двух видов: короткая (BRC1a S ) и длинная версия с альтернативным соединением (BRC1a L ). Оба приводят к образованию белков, но более короткая форма является цитоплазматической и не связывается с генами-мишенями в ядре.Соотношение между этими двумя формами меняется при обезглавливании побегов картофеля, нахождении в темноте и в условиях низкого R: FR (Nicolas et al., 2015). В то время как декапитация приводит к относительному увеличению BRC1a S , темнота и обработка с низким R: FR приводят к относительному увеличению содержания BRC1a L . Более длинный белок BRC1a L впоследствии ингибирует удлинение подмышечных ветвей в побегах и столонах картофеля (Nicolas et al., 2015). Arabidopsis brc1 и brc2 ( tcp12 и tcp18 ) демонстрируют пониженный ответ на R: FR, и ответ отсутствует у двойного мутанта brc1 brc2 (González-Grandío et al., 2013). Факторы транскрипции TCP также участвуют в разрастании пазушных зачатков петунии. Здесь Gh TCP3 действует вместе с DECREASED APICAL DOMINANCE 2, рецепторным белком, который обычно инактивирует стриголактоны в ответ на снижение R: FR (Drummond et al., 2015). Рис OsTCP15 участвует в удлинении мезокотилей в ответ на темноту и реагирует на лечение стриголактоном и цитокинином, подчеркивая взаимодействие между TCP и различными гормонами растений в регуляции развития, которая реагирует на окружающую среду (Hu et al., 2014).
Более длинный белок BRC1a L впоследствии ингибирует удлинение подмышечных ветвей в побегах и столонах картофеля (Nicolas et al., 2015). Arabidopsis brc1 и brc2 ( tcp12 и tcp18 ) демонстрируют пониженный ответ на R: FR, и ответ отсутствует у двойного мутанта brc1 brc2 (González-Grandío et al., 2013). Факторы транскрипции TCP также участвуют в разрастании пазушных зачатков петунии. Здесь Gh TCP3 действует вместе с DECREASED APICAL DOMINANCE 2, рецепторным белком, который обычно инактивирует стриголактоны в ответ на снижение R: FR (Drummond et al., 2015). Рис OsTCP15 участвует в удлинении мезокотилей в ответ на темноту и реагирует на лечение стриголактоном и цитокинином, подчеркивая взаимодействие между TCP и различными гормонами растений в регуляции развития, которая реагирует на окружающую среду (Hu et al., 2014).
Viola et al. (2013) показали, что TCP класса I содержат консервативный цистеин-20, который чувствителен к обработке оксидантами в зависимости от дозы. Это окислительно-восстановительное поведение TCP15 важно из-за его влияния на биосинтез антоцианов. Мутант, в котором цистеин-20 TCP15 был заменен серином, накапливает меньше антоциана при высоком световом стрессе, чем растения дикого типа (Viola et al., 2016). Экстракты растений со сверхэкспрессией TCP15 показали, что длительное пребывание в условиях высокой освещенности приводит к отмене ДНК-связывающей активности TCP15 in vivo , отражая фенотип in vitro (Viola et al., 2013, 2016). Таким образом, функция TCP15 реагирует на вход высокой освещенности. Хотя антоциановая реакция не является прямой реакцией развития, дальнейший анализ может показать, что существует эффект развития.
Это окислительно-восстановительное поведение TCP15 важно из-за его влияния на биосинтез антоцианов. Мутант, в котором цистеин-20 TCP15 был заменен серином, накапливает меньше антоциана при высоком световом стрессе, чем растения дикого типа (Viola et al., 2016). Экстракты растений со сверхэкспрессией TCP15 показали, что длительное пребывание в условиях высокой освещенности приводит к отмене ДНК-связывающей активности TCP15 in vivo , отражая фенотип in vitro (Viola et al., 2013, 2016). Таким образом, функция TCP15 реагирует на вход высокой освещенности. Хотя антоциановая реакция не является прямой реакцией развития, дальнейший анализ может показать, что существует эффект развития.
Не только свет влияет на TCP, но также воспринимаются и другие сигналы, которые приводят к регуляции TCP-опосредованного роста. Например, Guan et al. (2014) показали, что TCP20 участвует в питании корней Arabidopsis . В экспериментах с раздвоенным корнем дикий тип Arabidopsis развивает увеличенное количество боковых корней в среде, содержащей высокие концентрации нитратов (т. е.е., 5 мМ NO 3 —) и почти полное отсутствие боковых корней в среде, содержащей низкие концентрации нитратов (т.е. 0 мМ NO 3 —). Tcp20 Растения не проявляют этого поведения, указывая на то, что регуляция кормления корней находится под контролем TCP20 (Guan et al., 2014). Интересно, что TCP20 уровни транскрипта не находятся под контролем уровней нитратов, указывая тем самым, что TCP20 регулируется на уровне белка, либо за счет образования специфических белок-белковых димеров в случае дефицита нитрата, либо через другой регуляторный механизм.У риса транскрипт класса I TCP OsTCP19 активируется во время солевого стресса и лечения дефицита воды (Mukhopadhyay et al., 2015). Гетерологичная сверхэкспрессия OsTCP19 в Arabidopsis приводит к уменьшению количества боковых корней, но к повышению устойчивости к абиотическому стрессу, т.е. растения лучше росли на среде, содержащей маннит, и лучше восстанавливались после обработки дефицитом воды. Здесь экспрессия LOX2 была снижена в сверхэкспрессорах OsTCP19, а гены передачи сигналов ABA были усилены (Mukhopadhyay et al., 2015). Недавние эксперименты выявили повышающую и понижающую регуляцию нескольких TCP в Arabidopsis при осмотическом стрессе, хотя функциональный анализ их роли в ответ на осмотический стресс еще не проводился (Kumar et al., 2016). Таким образом, эти несколько результатов являются первым признаком того, что TCP не являются просто статическими регуляторами развития, но они действительно транслируют сигналы окружающей среды в регуляцию роста (рис. 4).
Здесь экспрессия LOX2 была снижена в сверхэкспрессорах OsTCP19, а гены передачи сигналов ABA были усилены (Mukhopadhyay et al., 2015). Недавние эксперименты выявили повышающую и понижающую регуляцию нескольких TCP в Arabidopsis при осмотическом стрессе, хотя функциональный анализ их роли в ответ на осмотический стресс еще не проводился (Kumar et al., 2016). Таким образом, эти несколько результатов являются первым признаком того, что TCP не являются просто статическими регуляторами развития, но они действительно транслируют сигналы окружающей среды в регуляцию роста (рис. 4).
РИСУНОК 4. Схематический рисунок, изображающий разнообразие сигналов окружающей среды, которые влияют на функции TCP в растениях (Mukhtar et al., 2011; Sugio et al., 2011; Balsemão-Pires et al., 2013; Гонсалес-Грандио и др., 2013; Нива и др., 2013; Виола и др., 2013, 2016; Guan et al., 2014; Hu et al., 2014; Ким и др., 2014; Mukhopadhyay et al., 2015; Николас и др. , 2015; Кумар и др., 2016).
, 2015; Кумар и др., 2016).
Заключение и прогноз
Факторы транскрипцииTCP играют роль во множестве процессов роста у широкого круга видов растений (рис. 1). Они влияют на рост напрямую через клеточный цикл и косвенно, влияя на гормональную сигнализацию растений и циркадные часы (рисунки 2 и 3).Кроме того, недавние открытия связывают контролируемые TCP реакции роста с такими сигналами окружающей среды, как R: FR, высокий световой стресс, солевой стресс или наличие или отсутствие питательных веществ.
Факторы транскрипции TCP участвуют во многих важных процессах развития и взаимодействуют с таким количеством гормонов растений, что вполне вероятно, что будущие исследования растений также откроют гораздо больше сигналов, на которые реагируют TCP. Это также будет означать, что будущие исследования TCP должны будут более подробно выяснить, как регулируется взаимодействие TCP с различными сетями сигнализации, чтобы обеспечить взвешенный ответ на экологические проблемы. Это исследование должно будет раскрыть роль димеризации, транскрипционной и посттранскрипционной регуляции, а также посттрансляционных модификаций в контроле и обеспечении определенных функций TCP в развитии растений.
Это исследование должно будет раскрыть роль димеризации, транскрипционной и посттранскрипционной регуляции, а также посттрансляционных модификаций в контроле и обеспечении определенных функций TCP в развитии растений.
Патогены растений нацелены на факторы транскрипции TCP, чтобы манипулировать архитектурой растений в свою пользу. Если патогены растений используют TCP в своих интересах, может быть, и нам следует. Факторы транскрипции TCP будут ценными инструментами в оптимизации архитектуры растений и укреплении растений в ответ на экологические проблемы.
Авторские взносы
SD подготовил, написал и критически отредактировал статью.
Финансирование
Работа в лаборатории автора поддерживается Фондом молодых исследователей Билефельда и основным грантом Университета Билефельда Д. Штайгеру.
Заявление о конфликте интересов
Автор заявляет, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Мы признательны Deutsche Forschungsgemeinschaft и Фонду публикаций открытого доступа Билефельдского университета за поддержку платы за обработку статей.
Сокращения
bHLH, основная спираль-петля-спираль; BRC, РАЗВЕТВЛЕННЫЙ; CCA1, CIRCADIAN CLOCK ASSOCIATED 1; CIN, CINCINNATA; CUC, ЧАШЕЧНЫЙ КОТИЛЕДОН; CYC, CYCLOIDEA; FT, ЦВЕТУЩИЙ ЛОКУС T; GA, гибберелловая кислота; ICS1, ИЗОХОРИСМАТНЫЙ СИНТАЗА 1; IDR, внутренне неупорядоченная область; челюсть -D, зазубренно-волнистая -D; ЛА, ЛАНЦЕОЛАТ; LHY, ПОЗДНИЙ УДЛИНЕННЫЙ ГИПОКОТИЛ; LOX, ЛИПОКСИГЕНАЗА; NLS — сигнал ядерной локализации; PRR, ПСЕВДО РЕГУЛЯТОР ОТВЕТСТВЕННОСТИ; TB1, ТЕОСИН РАЗВЕТВЛЕННЫЙ 1; TCP, TEOSINTE BRANCHED 1, CYCLOIDEA, PCF1; TOC1, ВРЕМЯ ВЫДВИЖЕНИЯ КАБИНЫ1.
Список литературы
Аггарвал П., Гупта М. Д., Джозеф А. П., Чаттерджи Н., Сринивасан Н. и Нат У. (2010). Идентификация специфических остатков связывания ДНК в семействе факторов транскрипции TCP Arabidopsis . Растительная клетка 22, 1174–1189. DOI: 10.1105 / tpc.109.066647
Растительная клетка 22, 1174–1189. DOI: 10.1105 / tpc.109.066647
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Агилар-Мартинес, Х.А., Поза-Каррион, К., и Кубас, П. (2007). Arabidopsis BRANCHED1 действует как интегратор сигналов ветвления в подмышечных зачатках. Растительная клетка 19, 458–472. DOI: 10.1105 / tpc.106.048934
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Агилар-Мартинес, Х.А., Синха, Н. (2013). Анализ роли TCP-генов Arabidopsis class I AtTCP7, AtTCP8, AtTCP22 и AtTCP23 в развитии листьев. Фронт. Plant Sci. 4: 406. DOI: 10.3389 / fpls.2013.00406
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Альварес, Дж. П., Фурумизу, К., Эфрони И., Эшед Ю. и Боуман Дж. Л. (2016). Активное подавление оркестровки меристемы листа определяет рост листа. eLife 5: e15023. DOI: 10.7554 / eLife.15023
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Andriankaja, M. E., Danisman, S., Mignolet-Spruyt, L. F., Claeys, H., Kochanke, I., Vermeersch, M., et al. (2014). Транскрипционная координация между дифференцировкой листовых клеток и развитием хлоропластов, установленная TCP20 и факторами транскрипции bHLH подгруппы Ib. Завод Мол. Биол. 85, 233–245. DOI: 10.1007 / s11103-014-0180-2
E., Danisman, S., Mignolet-Spruyt, L. F., Claeys, H., Kochanke, I., Vermeersch, M., et al. (2014). Транскрипционная координация между дифференцировкой листовых клеток и развитием хлоропластов, установленная TCP20 и факторами транскрипции bHLH подгруппы Ib. Завод Мол. Биол. 85, 233–245. DOI: 10.1007 / s11103-014-0180-2
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бай Ф., Райнхеймер Р., Дюрантини Д., Келлог Э. А. и Шмидт Р. Дж. (2012). Фактор транскрипции TCP, BRANCH ANGLE DEFECTIVE 1 (BAD1), необходим для нормального образования угла ветвления у кукурузы. Proc. Natl. Акад. Sci. США 109, 12225–12230. DOI: 10.1073 / pnas.1202439109
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Баллестер, П., Наваррете-Гомес, М., Карбонеро, П., Оньяте-Санчес, Л., и Феррандис, К. (2015). Распространение листьев у Arabidopsis контролируется регуляторным модулем TCP-NGA, вероятно, консервативным у отдаленно родственных видов. Physiol. Растение. 155, 21–32. DOI: 10.1111 / ppl.12327
Physiol. Растение. 155, 21–32. DOI: 10.1111 / ppl.12327
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Balsemão-Pires, E., Andrade, L.R., and Sachetto-Martins, G. (2013). Функциональное исследование TCP23 в Arabidopsis thaliana в процессе развития растений. Plant Physiol. Биохим. 67, 120–125. DOI: 10.1016 / j.plaphy.2013.03.009
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бен-Гера, Х., Ори, Н. (2012). Ауксин и ЛАНЦЕОЛАТ влияют на форму листьев томата через различные процессы развития. Завод Сигнал. Behav. 7, 1255–1257. DOI: 10.4161 / psb.21550
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бергер, Б.А., Томпсон, В., Лим, А., Ричильяно, В., и Ховарт, Д.Г. (2016). Выявление двусторонней симметрии Knautia macedonica capitula связано с изменениями экспрессии вентральных лепестков генов, подобных CYCLOIDEA. EvoDevo 7, 8. DOI: 10.1186 / s13227-016-0045-7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Браун, Н. , Жермен, А., де, С., Пилло, Ж.-П., Буте-Мерси, С., Далмайс, М., и др. (2012). Фактор транскрипции TCP гороха PsBRC1 действует ниже стриголактонов, контролируя ветвление побегов. Plant Physiol. 158, 225–238. DOI: 10.1104 / стр.111.182725
, Жермен, А., де, С., Пилло, Ж.-П., Буте-Мерси, С., Далмайс, М., и др. (2012). Фактор транскрипции TCP гороха PsBRC1 действует ниже стриголактонов, контролируя ветвление побегов. Plant Physiol. 158, 225–238. DOI: 10.1104 / стр.111.182725
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Брохольм, С. К., Тяхтихарью, С., Лайтинен, Р. А. Э., Альберт, В. А., Тери, Т. Х., и Эломаа, П. (2008). Фактор транскрипции домена TCP контролирует спецификацию типа цветка вдоль радиальной оси соцветия Gerbera (Asteraceae). Proc. Natl. Акад. Sci. 105, 9117–9122. DOI: 10.1073 / pnas.0801359105
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бурко, Ю., Шлейзер-Бурко, С., Янаи, О., Шварц, И., Зельник, И. Д., Якоб-Хирш, Дж. И др. (2013). Роль факторов транскрипции APETALA1 / FRUITFULL в развитии листьев томата. Растительная клетка 25, 2070–2083. DOI: 10.1105 / tpc.113.113035
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Busch, A. , Horn, S., Mühlhausen, A., Mummenhoff, K., and Zachgo, S. (2012). Моносимметрия венчика: эволюция морфологической новизны в семействе brassicaceae. Мол. Биол.Evol. 29, 1241–1254. DOI: 10.1093 / molbev / msr297
, Horn, S., Mühlhausen, A., Mummenhoff, K., and Zachgo, S. (2012). Моносимметрия венчика: эволюция морфологической новизны в семействе brassicaceae. Мол. Биол.Evol. 29, 1241–1254. DOI: 10.1093 / molbev / msr297
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чалла, К. Р., Аггарвал, П., и Нат, У. (2016). Активация YUCCA5 фактором транскрипции TCP4 объединяет онтогенетические и экологические сигналы, способствующие удлинению гипокотиля у Arabidopsis . Растительная клетка 28, 2117–2130. DOI: 10.1105 / TPC.16.00360
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Claßen-Bockhoff, R., Руонала, Р., Булл-Хереню, К., Марчант, Н., и Альберт, В.А. (2013). Уникальный псевдантий Actinodium (Myrtaceae) — морфологическое повторное исследование и возможная регуляция генами, подобными CYCLOIDEA. EvoDevo 4, 8. DOI: 10.1186 / 2041-9139-4-8
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Корли, С. Б., Карпентер, Р., Копси, Л., и Коэн, Э. (2005). Цветочная асимметрия включает взаимодействие между факторами транскрипции TCP и MYB в Antirrhinum . Proc. Natl. Акад. Sci. США 102, 5068–5073. DOI: 10.1073 / pnas.0501340102
Б., Карпентер, Р., Копси, Л., и Коэн, Э. (2005). Цветочная асимметрия включает взаимодействие между факторами транскрипции TCP и MYB в Antirrhinum . Proc. Natl. Акад. Sci. США 102, 5068–5073. DOI: 10.1073 / pnas.0501340102
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Коста, М. М., Фокс, С., Ханна, А. И., Бакстер, К., и Коэн, Э. (2005). Эволюция регуляторных взаимодействий, контролирующих асимметрию цветков. Разработка 132, 5093–5101. DOI: 10.1242 / dev.02085
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кубас П., Лаутер Н., Добли Дж. И Коэн Э.(1999a). Домен TCP: мотив, обнаруженный в белках, регулирующих рост и развитие растений. Plant J. 18, 215–222. DOI: 10.1046 / j.1365-313X.1999.00444.x
CrossRef Полный текст | Google Scholar
Данисман, С., Дейк, А. Д. Дж., Ван Бимбо, А., Вал, Ф., ван дер Хенниг, Л., Фолтер, С., и др. (2013). Анализ функциональной избыточности в семействе факторов транскрипции Arabidopsis TCP.![]() J. Exp. Бот. 64, 5673–5685. DOI: 10.1093 / jxb / ert337
J. Exp. Бот. 64, 5673–5685. DOI: 10.1093 / jxb / ert337
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Данисман, С., Wal, F., van der Dhondt, S., Waites, R., Folter, S., de Bimbo, A., et al. (2012). Arabidopsis Факторы транскрипции TCP класса I и класса II антагонистически регулируют метаболизм жасмоновой кислоты и развитие листьев. Plant Physiol. 159, 1511–1523. DOI: 10.1104 / стр.112.200303
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дарвин, К. Р. (1868). Разнообразие животных и растений при одомашнивании. Лондон: Джон Мюррей.
Google Scholar
Дас Гупта, М., Аггарвал П., Нат У. (2014). CINCINNATA в Antirrhinum majus непосредственно модулирует гены, участвующие в передаче сигналов цитокинина и ауксина. New Phytol. 204, 901–912. DOI: 10.1111 / Nph.12963
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Davière, J.-M. , Wild, M., Regnault, T., Baumberger, N., Eisler, H., Genschik, P., et al. (2014). Взаимодействия TCP-DELLA класса I на верхушке побега соцветия определяют высоту растения. Curr. Биол. 24, 1923–1928.DOI: 10.1016 / j.cub.2014.07.012
, Wild, M., Regnault, T., Baumberger, N., Eisler, H., Genschik, P., et al. (2014). Взаимодействия TCP-DELLA класса I на верхушке побега соцветия определяют высоту растения. Curr. Биол. 24, 1923–1928.DOI: 10.1016 / j.cub.2014.07.012
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Dereeper, A., Guignon, V., Blanc, G., Audic, S., Buffet, S., Chevenet, F., et al. (2008). Phylogeny.fr: надежный филогенетический анализ для неспециалистов. Nucleic Acids Res. 36, W465 – W469. DOI: 10.1093 / nar / gkn180
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Doebley, J., Stec, A., and Gustus, C. (1995). teosinte разветвленный1 и происхождение кукурузы: свидетельства эпистаза и эволюции доминирования. Генетика 141, 333–346.
PubMed Аннотация | Google Scholar
Дорнелас, М. К., Патрез, К. М., Ангенент, Г. К., и Имминк, Р. Г. Х. (2011). MADS: недостающее звено между идентичностью и ростом? Trends Plant Sci. 16, 89–97. DOI: 10.1016 / j.tplants.2010.11.003
16, 89–97. DOI: 10.1016 / j.tplants.2010.11.003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Драммонд, Р. С. М., Янссен, Б. Дж., Луо, З., Оплаат, К., Леджер, С. Е., Волерс, М. В. и др. (2015). Экологический контроль разветвления Петуния . Plant Physiol. 168, 735–751. DOI: 10.1104 / стр.15.00486
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дан, Э. А., Жермен, А., де, С., Рамо, К., и Беверидж, К. А. (2012). Антагонистическое действие стриголактона и цитокинина в борьбе с отрастанием почек. Plant Physiol. 158, 487–498. DOI: 10.1104 / стр.111.186783
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Эфрони И., Блюм Э., Гольдшмидт А. и Эшед Ю.(2008). Длительный и динамичный график созревания лежит в основе развития листьев Arabidopsis . Растительная клетка 20, 2293–2306. DOI: 10.1105 / tpc.107.057521
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Эфрони И. , Хан С.-К., Ким Х. Дж., Ву М.-Ф., Штайнер Э., Бирнбаум К. Д. и др. (2013). Регулирование созревания листьев посредством модуляции цитокининовых ответов, опосредованной хроматином. Dev. Cell 24, 438–445. DOI: 10.1016 / j.devcel.2013.01.019
, Хан С.-К., Ким Х. Дж., Ву М.-Ф., Штайнер Э., Бирнбаум К. Д. и др. (2013). Регулирование созревания листьев посредством модуляции цитокининовых ответов, опосредованной хроматином. Dev. Cell 24, 438–445. DOI: 10.1016 / j.devcel.2013.01.019
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фамбрини М., Сальвини М. и Пуглиси К. (2012). Опосредованная транспозоном инактивация гена, подобного CYCLOIDEA, приводит к появлению полисимметричных и андрогинных лучевых цветков у Helianthus annuus . Genetica 139, 1521–1529. DOI: 10.1007 / s10709-012-9652-y
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Жиро, Э., Нг, С., Кэрри, К., Дункан, О., Лоу, Дж., Ли, К.P., et al. (2010). Факторы транскрипции TCP связывают регуляцию генов, кодирующих митохондриальные белки, с циркадными часами Arabidopsis thaliana . Растительная клетка 22, 3921–3934. DOI: 10.1105 / tpc.110.074518
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гонсалес-Грандио, Э. , Поза-Каррион, К., Сорзано, К. О. С., и Кубас, П. (2013). BRANCHED1 способствует покою пазушных почек в ответ на тень у Arabidopsis . Растительная клетка 25, 834–850.DOI: 10.1105 / tpc.112.108480
, Поза-Каррион, К., Сорзано, К. О. С., и Кубас, П. (2013). BRANCHED1 способствует покою пазушных почек в ответ на тень у Arabidopsis . Растительная клетка 25, 834–850.DOI: 10.1105 / tpc.112.108480
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гуань, П., Ван, Р., Накри, П., Бретон, Г., Кей, С. А., Прунеда-Пас, Дж. Л. и др. (2014). Сбор нитратов корнями Arabidopsis опосредуется транскрипционным фактором TCP20 через системный сигнальный путь. Proc. Natl. Акад. Sci. США 111, 15267–15272. DOI: 10.1073 / pnas.1411375111
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Го, З., Фудзиока, С., Бланкафлор, Э. Б., Мяо, С., Гоу, X., и Ли, Дж. (2010). TCP1 Модулирует биосинтез брассиностероидов, регулируя экспрессию ключевого биосинтетического гена DWARF4 в Arabidopsis thaliana . Растительная клетка 22, 1161–1173. DOI: 10.1105 / tpc.109.069203
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Hao, J. , Tu, L., Hu, H., Tan, J., Deng, F., Tang, W., et al. (2012). GbTCP, фактор транскрипции TCP хлопка, обеспечивает удлинение волокон и развитие корневых волосков с помощью сложной регулирующей системы. J. Exp. Бот. 63, 6267–6281. DOI: 10.1093 / jxb / ers278
, Tu, L., Hu, H., Tan, J., Deng, F., Tang, W., et al. (2012). GbTCP, фактор транскрипции TCP хлопка, обеспечивает удлинение волокон и развитие корневых волосков с помощью сложной регулирующей системы. J. Exp. Бот. 63, 6267–6281. DOI: 10.1093 / jxb / ers278
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хорн С., Пабон-Мора Н., Тойс В. С., Буш А. и Закго С. (2015). Анализ класса CYC / TB1 факторов транскрипции TCP у базальных покрытосеменных и магнолиевых. Plant J. 81, 559–571. DOI: 10.1111 / tpj.12750
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ховарт, Д. Г., и Донохью, М. Дж. (2006).Филогенетический анализ клады «ECE» (CYC / TB1) выявляет дупликации, предшествовавшие основным эвдикотам. Proc. Natl. Акад. Sci. США 103, 9101–9106. DOI: 10.1073 / pnas.0602827103
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ховарт, Д. Г., Мартинс, Т., Чимни, Э., и Донохью, М. Дж. (2011). Диверсификация экспрессии CYCLOIDEA в эволюции билатеральной симметрии цветков у Caprifoliaceae и Lonicera (Dipsacales). Ann.Бот. 107, 1521–1532. DOI: 10.1093 / aob / mcr049
(2011). Диверсификация экспрессии CYCLOIDEA в эволюции билатеральной симметрии цветков у Caprifoliaceae и Lonicera (Dipsacales). Ann.Бот. 107, 1521–1532. DOI: 10.1093 / aob / mcr049
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ху, З., Ямаути, Т., Янг, Дж., Джикумару, Ю., Цучида-Маяма, Т., Итикава, Х., и др. (2014). Стриголактон и цитокинин антагонистически регулируют удлинение мезокотилей риса в темноте. Physiol растительных клеток. 55, 30–41. DOI: 10.1093 / pcp / pct150
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Яник К., Митхёфер А., Раффайнер, М., Штельмах, Х., Хауз, Б., и Шлинк, К. (2016). Эффектор фитоплазмы пролиферации яблони нацелен на факторы транскрипции TCP — обобщенная стратегия вирулентности фитоплазмы? Мол. Завод Патол. doi: 10.1111 / mpp.12409 [Epub перед печатью].
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Juntheikki-Palovaara, I. , Tähtiharju, S., Lan, T., Broholm, S.K., Rijpkema, A.S., Ruonala, R., et al. (2014). Функциональная диверсификация дублированных генов клады CYC2 в регуляции развития соцветий у Gerbera hybrida (Asteraceae). Plant J. 79, 783–796. DOI: 10.1111 / tpj.12583
, Tähtiharju, S., Lan, T., Broholm, S.K., Rijpkema, A.S., Ruonala, R., et al. (2014). Функциональная диверсификация дублированных генов клады CYC2 в регуляции развития соцветий у Gerbera hybrida (Asteraceae). Plant J. 79, 783–796. DOI: 10.1111 / tpj.12583
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кауфманн, К., Муиньо, Дж. М., Хорегуи, Р., Аирольди, К. А., Смачняк, К., Краевски, П. и др. (2009). Гены-мишени транскрипционного фактора MADS SEPALLATA3: интеграция онтогенетических и гормональных путей в цветке Arabidopsis . PLoS Biol. 7: e1000090. DOI: 10.1371 / journal.pbio.1000090
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кауфманн, К., Wellmer, F., Muiño, J.M., Ferrier, T., Wuest, S.E., Kumar, V., et al. (2010). Аранжировка цветочной инициации от APETALA1. Science 328, 85–89. DOI: 10.1126 / science.1185244
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кибл, Ф.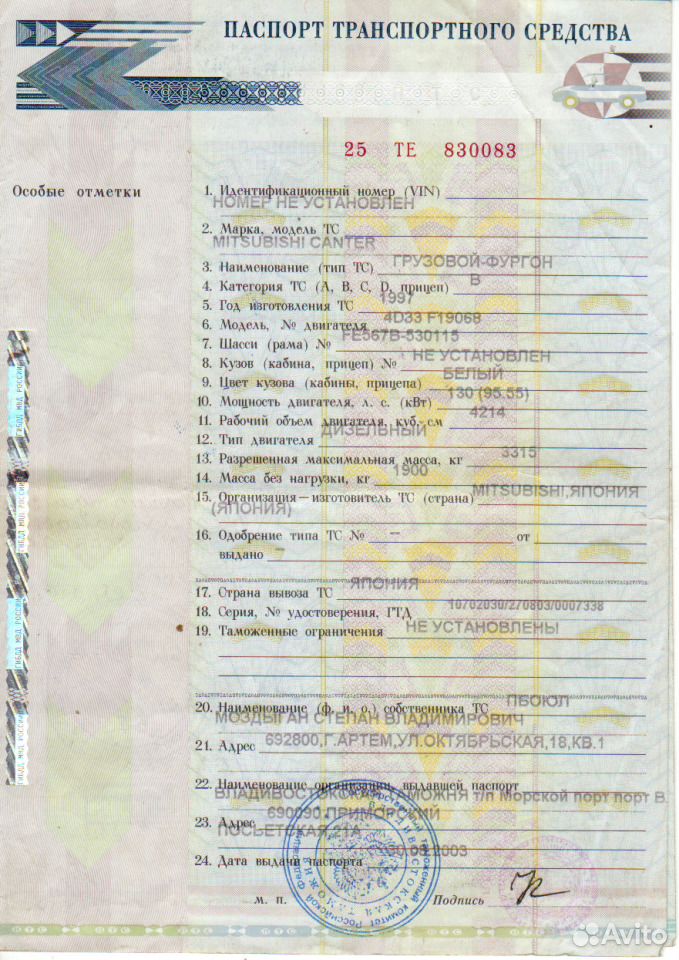 , Пелью, М. К., и Джонс, В. Н. (1910). Наследование Op peloria и цветочная окраска наперстянки (наперстянка пурпурная). New Phytol. 9, 68–77. DOI: 10.1111 / j.1469-8137.1910.tb05554.x
, Пелью, М. К., и Джонс, В. Н. (1910). Наследование Op peloria и цветочная окраска наперстянки (наперстянка пурпурная). New Phytol. 9, 68–77. DOI: 10.1111 / j.1469-8137.1910.tb05554.x
CrossRef Полный текст | Google Scholar
Киффер, М., Мастер, В., Уэйтс, Р., и Дэвис, Б. (2011). TCP14 и TCP15 влияют на длину междоузлий и форму листьев у Arabidopsis . Plant J. 68, 147–158. DOI: 10.1111 / j.1365-313X.2011.04674.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ким, М., Цуй, М.-Л., Кубас, П., Гиллис, А., Ли, К., Чепмен, М.А., и др. (2008). Регуляторные гены контролируют ключевой морфологический и экологический признак, передаваемый между видами. Наука 322, 1116–1119.DOI: 10.1126 / science.1164371
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ким, С. Х., Сон, Г. Х., Бхаттачарджи, С., Ким, Х. Дж., Нам, Дж. К., Нгуен, П. Д. Т. и др. (2014). Иммунный адаптер SRFR1 арабидопсиса Arabidopsis взаимодействует с факторами транскрипции TCP, которые вносят избыточный вклад в иммунитет, запускаемый эффектором. Plant J. 78, 978–989. DOI: 10.1111 / tpj.12527
Plant J. 78, 978–989. DOI: 10.1111 / tpj.12527
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Косуги, С.и Охаши Ю. (1997). PCF1 и PCF2 специфически связываются с цис-элементами в гене ядерного антигена пролиферирующих клеток риса. Растительная клетка 9, 1607–1619. DOI: 10.1105 / tpc.9.9.1607
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Косуги, С., Сузука, И., и Охаши, Ю. (1995). Два из трех промоторных элементов, идентифицированных в гене риса для ядерного антигена пролиферирующих клеток, важны для меристематической тканеспецифической экспрессии. Plant J. 7, 877–886.DOI: 10.1046 / j.1365-313X.1995.07060877.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кояма Т., Фурутани М., Тасака М. и Оме-Такаги М. (2007). Факторы транскрипции TCP контролируют морфологию боковых органов побегов посредством отрицательной регуляции экспрессии гранично-специфических генов у Arabidopsis . Растительная клетка 19, 473–484. DOI: 10.1105 / tpc.106.044792
Растительная клетка 19, 473–484. DOI: 10.1105 / tpc.106.044792
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кояма Т., Мицуда, Н., Секи, М., Шинозаки, К., и Оме-Такаги, М. (2010a). Факторы транскрипции TCP регулируют активность ASYMMETRIC LEAVES1 и miR164, а также реакцию ауксина во время дифференцировки листьев у Arabidopsis . Растительная клетка 22, 3574–3588. DOI: 10.1105 / tpc.110.075598
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кояма Т., Сато Ф. и Оме-Такаги М. (2010b). Роль TCP1 в продольном удлинении листьев у Arabidopsis . Biosci. Biotechnol. Биохим. 74, 2145–2147. DOI: 10.1271 / bbb.100442
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кумар Д., Хазра С., Датта Р. и Чаттопадхьяй С. (2016). Анализ транскриптома мутантов Arabidopsis предполагает наличие перекрестных помех между ABA, этиленом и GSH в отношении комбинированного холодового и осмотического стресса. Sci. Реп. 6, 36867. doi: 10.1038 / srep36867
Sci. Реп. 6, 36867. doi: 10.1038 / srep36867
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Летунич, И., и Борк, П. (2016). Интерактивное древо жизни (iTOL) v3: онлайн-инструмент для отображения и аннотации филогенетических и других деревьев. Nucleic Acids Res. 44, W242 – W245. DOI: 10.1093 / nar / gkw290
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ли К., Потущак Т., Колон-Кармона А., Гутьеррес Р. А. и Доернер П. (2005). Arabidopsis TCP20 связывает регуляцию роста и пути контроля клеточного деления. Proc. Natl. Акад. Sci.США 102, 12978–12983. DOI: 10.1073 / pnas.0504039102
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Линней, К., и Рудберг, Д. (1744). Dissertatio Botanica de Peloria. Упсала: Q-Med AB.
Google Scholar
Лю Дж., Перумал Н. Б., Олдфилд К. Дж., Су, Э. У., Уверский В. Н. и Дункер А. К. (2006). Внутреннее нарушение факторов транскрипции. Биохимия (Моск.) 45, 6873–6888. DOI: 10.1021 / bi0602718
Биохимия (Моск.) 45, 6873–6888. DOI: 10.1021 / bi0602718
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лусеро, Л.Э., Уберти-Манассеро, Н. Г., Арсе, А. Л., Коломбатти, Ф., Алемано, С. Г., и Гонсалес, Д. Х. (2015). TCP15 модулирует ответы цитокининов и ауксинов во время развития гинецея у Arabidopsis . Plant J. 84, 267–282. DOI: 10.1111 / tpj.12992
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Луо Д., Карпентер Р., Винсент К., Копси Л. и Коэн Э. (1996). Происхождение цветочной асимметрии у Antirrhinum . Природа 383, 794–799.DOI: 10.1038 / 383794a0
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ма, X., Ма, Дж., Фань, Д., Ли, К., Цзян, Ю., и Луо, К. (2016). Полногеномная идентификация факторов транскрипции семейства TCP из populus euphratica и их участие в регуляции формы листа. Sci. Rep. 6: 32795. DOI: 10.1038 / srep32795
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мао, Ю. , Ву, Ф., Ю, X., Бай, Дж., Чжун, В., и Хэ, Ю.(2014). Нацеленный на microRNA319a Brassica rapa ssp. pekinensis TCP гены модулируют форму головы китайской капусты за счет дифференциальной остановки деления клеток в областях листа. Plant Physiol. 164, 710–720. DOI: 10.1104 / стр.113.228007
, Ву, Ф., Ю, X., Бай, Дж., Чжун, В., и Хэ, Ю.(2014). Нацеленный на microRNA319a Brassica rapa ssp. pekinensis TCP гены модулируют форму головы китайской капусты за счет дифференциальной остановки деления клеток в областях листа. Plant Physiol. 164, 710–720. DOI: 10.1104 / стр.113.228007
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мимида, Н., Кидо, С.-И., Иванами, Х., Мория, С., Абэ, К., Вугд, К., и др. (2011). Белки Apple FLOWERING LOCUS T взаимодействуют с факторами транскрипции, участвующими в росте клеток и развитии органов. Tree Physiol. 31, 555–566. DOI: 10.1093 / treephys / tpr028
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мизуно, С., Сонода, М., Тамура, Ю., Нишино, Э., Судзуки, Х., Сато, Т. и др. (2015). Локус Chiba Tendril-Less определяет идентичность усикового органа дыни ( Cucumis melo L.) и потенциально кодирует специфический для усиков гомолог TCP. J. Plant Res. 128, 941–951. DOI: 10.1007 / s10265-015-0747-2
128, 941–951. DOI: 10.1007 / s10265-015-0747-2
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мур, М., Прюфер, Н., Паулат, М., Тейхманн, Т. (2016). Нокдаун генов биосинтеза стриголактона у Populus влияет на экспрессию BRANCHED1 и архитектуру побегов. New Phytol. 212, 613–626. DOI: 10.1111 / nph.14076
CrossRef Полный текст
Мухопадхьяй П., Тяги А. К., Тяги А. К. (2015). OsTCP19 влияет на передачу сигналов онтогенетического и абиотического стресса, модулируя пути, опосредованные ABI4. Sci. Реп. 5: 9998. DOI: 10.1038 / srep09998
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мухтар, М.S., Carvunis, A.-R., Dreze, M., Epple, P., Steinbrenner, J., Moore, J., et al. (2011). Независимо возникшие эффекторы вирулентности сходятся в узлах сети иммунной системы растений. Наука 333, 596–601. DOI: 10.1126 / science.1203659
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Наг, А. , Кинг, С., и Джек, Т. (2009). Нацеливание miR319a на TCP4 является критическим для роста и развития лепестков у Arabidopsis . Proc. Natl. Акад. Sci.США 106, 22534–22539. DOI: 10.1073 / pnas.0
, Кинг, С., и Джек, Т. (2009). Нацеливание miR319a на TCP4 является критическим для роста и развития лепестков у Arabidopsis . Proc. Natl. Акад. Sci.США 106, 22534–22539. DOI: 10.1073 / pnas.0
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Наво, О., Дабос, П., Карнус, Э., Тремусейг, Д., и Эрве, К. (2007). Факторы транскрипции TCP предшествуют появлению наземных растений. J. Mol. Evol. 65, 23–33. DOI: 10.1007 / s00239-006-0174-z
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Николас, М., Родригес-Буэй, М. Л., Франко-Соррилья, Дж. М., и Кубас, П.(2015). Недавно разработанный альтернативный сайт сплайсинга в гене BRANCHED1a контролирует архитектуру растения картофеля. Curr. Биол. 25, 1799–1809. DOI: 10.1016 / j.cub.2015.05.053
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Нива, М., Даймон, Ю., Куротани, К., Хиго, А., Прунэда-Пас, Дж. Л., Бретон, Г. , и др. (2013). BRANCHED1 взаимодействует с FLOWERING LOCUS T для подавления цветочного перехода пазушных меристем у Arabidopsis . Растительная клетка 25, 1228–1242.DOI: 10.1105 / TPC.112.109090
, и др. (2013). BRANCHED1 взаимодействует с FLOWERING LOCUS T для подавления цветочного перехода пазушных меристем у Arabidopsis . Растительная клетка 25, 1228–1242.DOI: 10.1105 / TPC.112.109090
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ори Н., Коэн А. Р., Эциони А., Бранд А., Янаи О., Шлейзер С. и др. (2007). Регуляция LANCEOLATE с помощью miR319 необходима для развития сложных листьев у томатов. Нат. Genet. 39, 787–791. DOI: 10.1038 / ng2036
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ортис-Рамирес, К., Эрнандес-Коронадо, М., Тамм, А., Катарино, Б., Ван, М., Долан Л. и др. (2016). Атлас транскриптомов Physcomitrella patens дает представление об эволюции и развитии наземных растений. Мол. Растение. 9, 205–220. DOI: 10.1016 / j.molp.2015.12.002
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Palatnik, J. F., Allen, E., Wu, X., Schommer, C., Schwab, R., Carrington, J.![]() C., et al. (2003). Контроль морфогенеза листа с помощью микроРНК. Nature 425, 257–263. DOI: 10.1038 / nature01958
C., et al. (2003). Контроль морфогенеза листа с помощью микроРНК. Nature 425, 257–263. DOI: 10.1038 / nature01958
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Поса-Каррион, К., Агилар-Мартинес, Дж. А., и Кубас, П. (2007). Роль TCP Gene BRANCHED1 в контроле ветвления побегов Arabidopsis . Завод Сигнал. Behav. 2, 551–552. DOI: 10.4161 / psb.2.6.4811
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Прунэда-Пас, Дж. Л., Бретон, Г., Нагель, Д. Х., Канг, С. Э., Боналди, К., Доэрти, К. Дж. И др. (2014). Ресурс в масштабе генома для функциональной характеристики факторов транскрипции Arabidopsis . Cell Rep. 8, 622–632. DOI: 10.1016 / j.celrep.2014.06.033
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Прунэда-Пас, Дж. Л., Бретон, Г., Пара, А., и Кей, С. А. (2009). Подход функциональной геномики показывает CHE как компонент циркадных часов Arabidopsis . Наука 323, 1481–1485. DOI: 10.1126 / science.1167206
Наука 323, 1481–1485. DOI: 10.1126 / science.1167206
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шоммер, К., Палатник, Дж.Ф., Аггарвал П., Шетелат А., Кубас П., Фармер Э. Э. и др. (2008). Контроль биосинтеза и старения жасмоната мишенями miR319. PLoS Biol. 6: e230. DOI: 10.1371 / journal.pbio.0060230
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шлейзер-Бурко, С., Бурко, Ю., Бен-Герцель, О., и Ори, Н. (2011). Программа динамического роста, регулируемая LANCEOLATE, позволяет гибко формировать листья. Развитие 138, 695–704. DOI: 10.1242 / dev.056770
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Смит, С.Дж. Э. (1821). Избранные переписки Линнея и других естествоиспытателей: из оригинальных рукописей . Харлоу: Лонгман.
Google Scholar
Штайнер, Э., Эфрони, И., Гопалрадж, М., Саатхофф, К., Ценг, Т.-С., Киффер, М., и др. (2012). O-связанная N-ацетилглюкозаминтрансфераза Arabidopsis SPINDLY взаимодействует с TCP класса I для облегчения цитокининовых ответов в листьях и цветках. Растительная клетка 24, 96–108. DOI: 10,1105 / тпк.111.093518
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сугио А., Королевство Х. Н., Маклин А. М., Грив В. М. и Хогенхаут С. А. (2011). Эффектор фитоплазменного белка SAP11 усиливает размножение насекомых-переносчиков путем манипулирования развитием растений и биосинтезом защитных гормонов. Proc. Natl. Акад. Sci. США 108, E1254 – E1263. DOI: 10.1073 / pnas.1105664108
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сугио, А., Маклин, А. М., Хогенхаут, С. А. (2014). Небольшой эффектор вирулентности фитоплазмы SAP11 содержит отдельные домены, необходимые для ядерного нацеливания и связывания и дестабилизации CIN-TCP. New Phytol. 202, 838–848. DOI: 10.1111 / Nph.12721
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сунь, Дж., Ци, Л., Ли, Ю., Чу, Дж., И Ли, К. (2012). PIF4-опосредованная активация экспрессии YUCCA8 интегрирует температуру в ауксиновый путь при регуляции роста гипокотилей Arabidopsis .![]() PLOS Genet 8: e1002594. DOI: 10.1371 / journal.pgen.1002594
PLOS Genet 8: e1002594. DOI: 10.1371 / journal.pgen.1002594
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Свёнтек, А., Ленджу, М., Ван Бокстаэле, Д., Инзе, Д., и Ван Онкелен, Х. (2002). Дифференциальный эффект жасмоновой кислоты и абсцизовой кислоты на развитие клеточного цикла в клетках табака BY-2. Plant Physiol. 128, 201–211. DOI: 10.1104 / pp.010592
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тяхтихарью, С., Rijpkema, A. S., Vetterli, A., Albert, V. A., Teeri, T. H., and Elomaa, P. (2012). Эволюция и диверсификация семейства генов CYC / TB1 у сложноцветных — сравнительное исследование на гербере (Mutisieae) и подсолнечнике (Heliantheae). Мол. Биол. Evol. 29, 1155–1166. DOI: 10.1093 / molbev / msr283
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Такеда Т., Сува Ю., Судзуки М., Китано Х., Уегучи-Танака М., Асикари М. и др. (2003). Ген OsTB1 отрицательно регулирует латеральное ветвление у риса. Plant J. 33, 513–520. DOI: 10.1046 / j.1365-313X.2003.01648.x
Plant J. 33, 513–520. DOI: 10.1046 / j.1365-313X.2003.01648.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тао, К., Го, Д., Вэй, Б., Чжан, Ф., Панг, К., Цзян, Х. и др. (2013). Репрессор транскрипции TIE1 связывает факторы транскрипции TCP с корепрессорами TOPLESS / TOPLESS-RELATED и модулирует развитие листа у Arabidopsis . Растительная клетка 25, 421–437. DOI: 10.1105 / tpc.113.109223
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Татэмацу, К., Накабаяси, К., Камия, Ю., и Намбара, Э. (2008). Фактор транскрипции AtTCP14 регулирует потенциал эмбрионального роста во время прорастания семян у Arabidopsis thaliana . Plant J. 53, 42–52. DOI: 10.1111 / j.1365-313X.2007.03308.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тиеулин-Пардо, Г., Авилан, Л., Кожадинович, М., и Гонтеро, Б. (2015). Сказочные «хвосты»: гибкость и функция внутренне неупорядоченных расширений в фотосинтетическом мире. Фронт. Мол. Biosci. 2:23. DOI: 10.3389 / fmolb.2015.00023
Фронт. Мол. Biosci. 2:23. DOI: 10.3389 / fmolb.2015.00023
CrossRef Полный текст | Google Scholar
Тремусайг, Д., Гарнье, Л., Барде, К., Дабос, П., Эрве, К., и Лескюр, Б. (2003). Внутренние теломерные повторы и сайты связывания белка «TCP-домен» взаимодействуют для регулирования экспрессии генов в циклических клетках Arabidopsis thaliana . Plant J. 33, 957–966. DOI: 10.1046 / j.1365-313X.2003.01682.x
CrossRef Полный текст | Google Scholar
Уберти-Манассеро, Н.Г., Лусеро, Л. Е., Виола, И. Л., Вегетти, А. К., и Гонсалес, Д. Х. (2012). Белок класса I AtTCP15 модулирует развитие растений посредством пути, который перекрывается с сигналом, на который влияют CIN-подобные белки TCP. J. Exp. Бот. 63, 809–823. DOI: 10.1093 / jxb / err305
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Valsecchi, I., Guittard-Crilat, E., Maldiney, R., Habricot, Y., Lignon, S., Lebrun, R., et al. (2013). Внутренне неупорядоченная С-концевая область транскрипционного фактора Arabidopsis thaliana TCP8 действует как домен трансактивации, так и самосборка. Мол. Биосист. 9, 2282–2295. DOI: 10.1039 / c3mb70128j
Мол. Биосист. 9, 2282–2295. DOI: 10.1039 / c3mb70128j
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Виола, И. Л., Камойрано, А., и Гонсалес, Д. Х. (2016). Редокс-зависимая модуляция биосинтеза антоцианов с помощью фактора транскрипции TCP TCP15 во время воздействия условий высокой интенсивности света у Arabidopsis . Plant Physiol. 170, 74–85. DOI: 10.1104 / стр.15.01016
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Виола, И.Л., Гюттлейн, Л. Н., Гонсалес, Д. Х. (2013). Редокс-модуляция регуляторов развития растений из семейства факторов транскрипции TCP I класса. Plant Physiol. 162, 1434–1447. DOI: 10.1104 / стр.113.216416
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Виола, И. Л., Райнхеймер, Р., Риполл, Р., Манассеро, Н. Г. У. и Гонсалес, Д. Х. (2012). Детерминанты специфичности связывания ДНК факторов транскрипции TCP I и II классов. Дж.Биол. Chem. 287, 347–356. DOI: 10.1074 / jbc.M111.256271
287, 347–356. DOI: 10.1074 / jbc.M111.256271
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Виола И. Л., Уберти Манассеро Н. Г., Риполл Р. и Гонсалес Д. Х. (2011). Фактор транскрипции TCP Arabidopsis класса I TCP AtTCP11 является регулятором развития с отчетливыми ДНК-связывающими свойствами из-за присутствия остатка треонина в положении 15 домена TCP. Biochem. J. 435, 143–155. DOI: 10.1042 / BJ20101019
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ван, Р.-Л., Стек, А., Эй, Дж., Люкенс, Л., и Добли, Дж. (1999). Пределы отбора при одомашнивании кукурузы. Природа 398, 236–239. DOI: 10.1038 / 18435
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Wang, S., Yang, X., Xu, M., Lin, X., Lin, T., Qi, J., et al. (2015). Редкий SNP идентифицировал фактор транскрипции TCP, необходимый для развития усиков огурца. Мол. Завод 8, 1795–1808. DOI: 10.1016 / j.molp.2015. 10.005
10.005
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ван, X., Гао, Дж., Чжу, З., Дун, X., Ван, X., Рен, Г., и др. (2015). Факторы транскрипции TCP имеют решающее значение для скоординированной регуляции экспрессии ISOCHORISMATE SYNTHASE 1 в Arabidopsis thaliana . Plant J. 82, 151–162. DOI: 10.1111 / tpj.12803
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Wang, Z., Luo, Y., Li, X., Wang, L., Xu, S., Yang, J., et al. (2008). Генетический контроль цветочной зигоморфии гороха ( Pisum sativum L.). Proc. Natl. Акад. Sci. США 105, 10414–10419. DOI: 10.1073 / pnas.08032
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вейр, И., Лу, Дж., Кук, Х., Каузье, Б., Шварц-Зоммер, З., и Дэвис, Б. (2004). CUPULIFORMIS устанавливает боковые границы органов в Antirrhinum . Развитие 131, 915–922. DOI: 10.1242 / dev.00993
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Велхен, Э. , Гонсалес, Д.Х. (2006). Избыточное количество элементов, распознаваемых факторами транскрипции TCP-домена, в вышестоящих областях ядерных генов, кодирующих компоненты механизма окислительного фосфорилирования митохондрий. Plant Physiol. 141, 540–545. DOI: 10.1104 / стр.105.075366
, Гонсалес, Д.Х. (2006). Избыточное количество элементов, распознаваемых факторами транскрипции TCP-домена, в вышестоящих областях ядерных генов, кодирующих компоненты механизма окислительного фосфорилирования митохондрий. Plant Physiol. 141, 540–545. DOI: 10.1104 / стр.105.075366
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Wu, J.-F., Tsai, H.-L., Joanito, I., Wu, Y.-C., Chang, C.-W., Li, Y.-H., et al. (2016). Комплекс LWD-TCP активирует утренний ген CCA1 в Arabidopsis . Нат. Commun. 7: 13181. DOI: 10.1038 / ncomms13181
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Янг, X., Чжао, X.-G., Li, C.-Q., Liu, J., Qiu, Z.-J., Dong, Y., et al. (2015). Отчетливые регуляторные изменения, лежащие в основе дифференциальной экспрессии генов TEOSINTE BRANCHED1-CYCLOIDEA-PROLIFERATING CELL FACTOR, связанные с вариациями лепестков зигоморфных цветков Petrocosmea spp. семейства Gesneriaceae. Plant Physiol. 169, 2138–2151.DOI: 10.1104 / стр.15.01181
169, 2138–2151.DOI: 10.1104 / стр.15.01181
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Юань, З., Гао, С., Сюэ, Д.-В., Луо, Д., Ли, Л.-Т., Дин, С.-Й., и др. (2009). RETARDED PALEA1 контролирует развитие палеа и цветочную зигоморфию риса. Plant Physiol. 149, 235–244. DOI: 10.1104 / стр.108.128231
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Zheng, X., Zhou, M., Yoo, H., Pruneda-Paz, J. L., Spivey, N. W., Kay, S.A., et al. (2015).Пространственная и временная регуляция биосинтеза растительного иммунного сигнала салициловой кислоты. Proc. Natl. Акад. Sci. США 112, 9166–9173. DOI: 10.1073 / pnas.1511182112
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Zhou, M., Li, D., Li, Z., Hu, Q., Yang, C., Zhu, L., et al. (2013). Конститутивная экспрессия гена miR319 изменяет развитие растений и повышает устойчивость к соли и засухе у трансгенных ползучих трав. Plant Physiol.