Боковой интервал: Безопасная дистанция и боковой интервал — Новости — ГИБДД отделения МВД — Государственные организации информируют
Дистанция и боковой интервал — Уроки вождения для начинающих
В различных странах мира есть конкретные рекомендации относительно безопасной дистанции до движущейся впереди машины. Все они сводятся к тому, что на сухой дороге безопасным будет расстояние, которое автомобиль пройдет за две секунды.
Иными словами, около той точки, мимо которой только что проехала движущаяся впереди машина, Вам советуют оказаться примерно через две секунды. В метрах это расстояние приблизительно равно величине, которая получается, если скорость, с которой движется Ваша машина, разделить на два (например, 60 км/ч : 2 = 30 м).
Рекомендации разумные. При любой скорости движения дистанция получается такой, которая должна обеспечить безопасную оставновку в случае, если впереди идущая машина вдруг резко затормозит. Но есть одна проблема. Если на наших дорогах только и делать, что увлеченно отсчитывать секунды, то дистанция-то будет неплохой, но как быть со всем остальным? Ведь нам надо искать знаки, спрятанные за деревьями и кустами, прогнозировать переключения сигналов светофора, выискивать глазами пешеходов, которые «мечтают» перебежать дорогу именно перед нашей машиной, объезжать ямы и колдобины, а также учитывать прочие особенности национальной езды.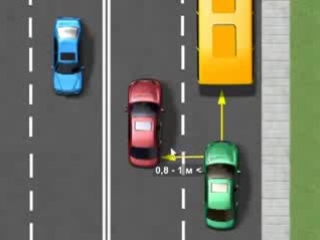
Поэтому вместо системы сухой математики советую Вам развивать у себя чувства времени, скорости и расстояния, применительно к конкретной ситуации. Поверьте, Вы в состоянии именно чувствовать безопасную дистанцию, для этого надо просто периодически задавать себе простой вопрос: «Какая у меня сейчас дистанция? Не слишком ли она мала или велика?» Причем это только на первое время. Позже достаточно будет лишь удерживать движущуюся впереди машину в поле своего периферического зрения, и оно Вам подскажет, оптимальная ли дистанция у Вас в данный момент.
На сухой дороге безопасной дистанцией до движущейся впереди машины считается расстояние, которое пройдет автомобиль примерно за две секунды.
А что делать, если в плотном потоке транспорта машина, движущаяся сзади, приблизилась к Вам на небезопасное расстояние?
Что ж, такое бывает. Водитель той машины хочет, чтобы Вы ехали быстрее или вообще освободили его полосу.
В таких случаях следует немного уменьшить скорость, но только очень плавно. Тем самым Вы увеличиваете дистанцию до впереди идущей машины, что позволит Вам применить в случае необходимости менее эффективное торможение, и водитель сзади идущей машины тоже сможет вовремя остановиться. А вообще стоит подумать, Ваша ли это полоса? В состоянии ли Вы поддерживать скорость и дистанцию, которые применяют большинство водителей при движении по этой полосе. Если Вы сомневаетесь, то лучше перестроиться на другую полосу.
Разумной должна быть не только дистанция, но и боковой интервал. Безопасным интервалом считается расстояние, приблизительно равное половине ширины автомобилф. Для среднестатического легкового автомобиля это: 1,7 м : 2 = 0,85 м, или 85 см. Такое расстояние позволяет водителю вовремя среагировать на внезапное изменение направления движения соседней или встречной машины.
Но есть опять проблемы. Всегда ли возможно поддерживать рекомендованный интервал? Надо ли его увеличивать пли уменьшать в зависимости от ситуации?
При выборе бокового интервала следует учитывать скорость и интенсивность движения, покрытие дороги и погодные условия, а также много других факторов.
При любом ухудшении дорожной ситуации боковой интервал необходимо увеличивать.
А если увеличивать некуда?
Тогда водитель должен следовать правилу: «Чем меньше боковой интервал, тем меньше скорость». Пытаясь проехать через узкую щель между припаркованными машинами, проезжая вплотную мимо стоящего на остановке автобуса и в других аналогичных случаях, когда боковой интервал слишком мал, скорость надо снижать вплоть до скорости движения пешехода. Если в транспортном потоке соседние автомобили зажимают Вас с боков, то и здесь есть выход — пропустите их вперед, для чего достаточно лишь немного снизить скорость.
вернуться к оглавлению «Уроки вождения»
- При увеличении скорости движения боковой интервал необходимо также увеличить.
- Чем меньше боковой интервал, тем меньше должна быть скорость движения.
ПДД РФ, 9.
 Расположение транспортных средств на проезжей части / КонсультантПлюс 9.1. Количество полос движения для безрельсовых транспортных средств определяется разметкой и (или) знаками 5.15.1, 5.15.2, 5.15.7, 5.15.8, а если их нет, то самими водителями с учетом ширины проезжей части, габаритов транспортных средств и необходимых интервалов между ними. При этом стороной, предназначенной для встречного движения на дорогах с двусторонним движением без разделительной полосы, считается половина ширины проезжей части, расположенная слева, не считая местных уширений проезжей части (переходно-скоростные полосы, дополнительные полосы на подъем, заездные карманы мест остановок маршрутных транспортных средств).(в ред. Постановлений Правительства РФ от 14.12.2005 N 767, от 10.05.2010 N 316)
Расположение транспортных средств на проезжей части / КонсультантПлюс 9.1. Количество полос движения для безрельсовых транспортных средств определяется разметкой и (или) знаками 5.15.1, 5.15.2, 5.15.7, 5.15.8, а если их нет, то самими водителями с учетом ширины проезжей части, габаритов транспортных средств и необходимых интервалов между ними. При этом стороной, предназначенной для встречного движения на дорогах с двусторонним движением без разделительной полосы, считается половина ширины проезжей части, расположенная слева, не считая местных уширений проезжей части (переходно-скоростные полосы, дополнительные полосы на подъем, заездные карманы мест остановок маршрутных транспортных средств).(в ред. Постановлений Правительства РФ от 14.12.2005 N 767, от 10.05.2010 N 316)редакции)9.1(1). На любых дорогах с двусторонним движением запрещается движение по полосе, предназначенной для встречного движения, если она отделена трамвайными путями, разделительной полосой, разметкой 1.
 1, 1.3 или разметкой 1.11, прерывистая линия которой расположена слева.9.2. На дорогах с двусторонним движением, имеющих четыре или более полосы, запрещается выезжать для обгона или объезда на полосу, предназначенную для встречного движения. На таких дорогах повороты налево или развороты могут выполняться на перекрестках и в других местах, где это не запрещено Правилами, знаками и (или) разметкой.
1, 1.3 или разметкой 1.11, прерывистая линия которой расположена слева.9.2. На дорогах с двусторонним движением, имеющих четыре или более полосы, запрещается выезжать для обгона или объезда на полосу, предназначенную для встречного движения. На таких дорогах повороты налево или развороты могут выполняться на перекрестках и в других местах, где это не запрещено Правилами, знаками и (или) разметкой.(см. текст в предыдущей редакции
)

(см. текст в предыдущей редакции
)
(см. текст в предыдущей редакции
)
 Выезд на левую полосу дорог с односторонним движением для остановки и стоянки осуществляется в соответствии с пунктом 12.1 Правил.(в ред. Постановлений Правительства РФ от 25.09.2003 N 595, от 10.05.2010 N 316)
Выезд на левую полосу дорог с односторонним движением для остановки и стоянки осуществляется в соответствии с пунктом 12.1 Правил.(в ред. Постановлений Правительства РФ от 25.09.2003 N 595, от 10.05.2010 N 316)(см. текст в предыдущей редакции
)
(см. текст в предыдущей редакции
)
9.5. Транспортные средства, скорость движения которых не должна превышать 40 км/ч или которые по техническим причинам не могут развивать такую скорость, должны двигаться по крайней правой полосе, кроме случаев объезда, обгона или перестроения перед поворотом налево разворотом или остановкой в разрешенных случаях на левой стороне дороги.
(см. текст в предыдущей редакции
)
 5 Правил. При этом не должно создаваться помех трамваю. Выезжать на трамвайные пути встречного направления запрещается. Если перед перекрестком установлены дорожные знаки 5.15.1 или 5.15.2, движение по трамвайным путям через перекресток запрещается.(в ред. Постановлений Правительства РФ от 24.01.2001 N 67, от 14.12.2005 N 767, от 28.03.2012 N 254)
5 Правил. При этом не должно создаваться помех трамваю. Выезжать на трамвайные пути встречного направления запрещается. Если перед перекрестком установлены дорожные знаки 5.15.1 или 5.15.2, движение по трамвайным путям через перекресток запрещается.(в ред. Постановлений Правительства РФ от 24.01.2001 N 67, от 14.12.2005 N 767, от 28.03.2012 N 254)(см. текст в предыдущей редакции
)
9.7. Если проезжая часть разделена на полосы линиями разметки, движение транспортных средств должно осуществляться строго по обозначенным полосам. Наезжать на прерывистые линии разметки разрешается лишь при перестроении.
9.8. При повороте на дорогу с реверсивным движением водитель должен вести транспортное средство таким образом, чтобы при выезде с пересечения проезжих частей транспортное средство заняло крайнюю правую полосу. Перестроение разрешается только после того, как водитель убедится, что движение в данном направлении разрешается и по другим полосам.
редакции)
9. 10. Водитель должен соблюдать такую дистанцию до движущегося впереди транспортного средства, которая позволила бы избежать столкновения, а также необходимый боковой интервал, обеспечивающий безопасность движения.
10. Водитель должен соблюдать такую дистанцию до движущегося впереди транспортного средства, которая позволила бы избежать столкновения, а также необходимый боковой интервал, обеспечивающий безопасность движения.
9.11. Вне населенных пунктов на дорогах с двусторонним движением, имеющих две полосы, водитель транспортного средства, для которого установлено ограничение скорости, а также водитель транспортного средства (состава транспортных средств) длиной более 7 м должен поддерживать между своим и движущимся впереди транспортным средством такую дистанцию, чтобы обгоняющие его транспортные средства могли без помех перестроиться на ранее занимаемую ими полосу. Это требование не действует при движении по участкам дорог, на которых запрещается обгон, а также при интенсивном движении и движении в организованной транспортной колонне.
(см. текст в предыдущей редакции
)
9.12. На дорогах с двусторонним движением при отсутствии разделительной полосы островки безопасности, тумбы и элементы дорожных сооружений (опоры мостов, путепроводов и тому подобное), находящиеся на середине проезжей части, водитель должен объезжать справа, если знаки и разметка не предписывают иное.
Открыть полный текст документа
ПДД 9.10 — Соблюдать дистанцию. Боковой интервал
Водитель должен соблюдать такую дистанцию до движущегося впереди транспортного средства, которая позволила бы избежать столкновения, а также необходимый боковой интервал, обеспечивающий безопасность движения.
В каких случаях следует увеличить боковой интервал?
| 1. | При разъезде с длинномерным транспортным средством. | |
| 2. | При движении по мокрому, скользкому или неровному покрытию. | |
| 3. | При встречном разъезде на большой скорости. | |
| 4. | Во всех перечисленных случаях. |
Ответ.
- Чем выше скорость при встречном разъезде транспортных средств, тем больше должен быть боковой интервал между ними для исключения возможного столкновения при неожиданном отклонении от траектории движения.

- При разъезде с длинномерным транспортным средством необходимо иметь запас бокового интервала во избежание столкновения с прицепом (полуприцепом), который смещается к центру поворота при выполнении различных маневров.
- При движении по мокрому, скользкому или неровному покрытию увеличение бокового интервала требуется во избежание столкновения, возникающего при потере устойчивости транспортного средства в неблагоприятных дорожных условиях.
Правильный ответ — во всех перечисленных случаях.
Укажите расстояние, под которым в Правилах подразумевается дистанция:
| 1. | Только А. | |
| 2. | Только Б. | |
| 3. | Только В. | |
| 4. | А и В. |
Под дистанцией Правила подразумевают расстояние (Б) до движущегося впереди ТС. Расстояние между попутными (В) и попутным и встречным автомобилями (А) называется боковым интервалом.
Зависит ли выбор бокового интервала от скорости движения?
| 1. | Выбор бокового интервала от скорости движения не зависит. | |
| 2. | При увеличении скорости движения боковой интервал необходимо увеличить. |
При увеличении скорости увеличиваются и поперечные отклонения от задаваемой водителем траектории движения. Это происходит за счет боковой эластичности колес, неровностей покрытия, воздействия ветра и люфтов в механизмах рулевого управления. Поэтому, увеличивая скорость, водители должны увеличивать боковой интервал.
Безопасной дистанцией при движении по сухой дороге на легковом автомобиле можно считать расстояние, которое автомобиль пройдет не менее чем за:
| 1. | 1 секунду. | |
| 2. | 2 секунды. | |
| 3. | 3 секунды. |
Безопасной дистанцией считается расстояние, которое может проехать автомобиль за время реакции водителя и время срабатывания тормозной системы.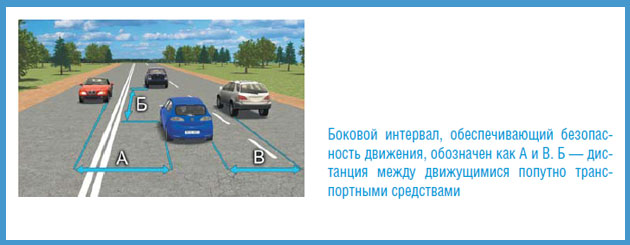 Время реакции водителя может колебаться от 0,4 до 1,6 секунд, время срабатывания тормозной системы с гидравлическим приводом может составлять от 0,1 до 0,4 секунд. То есть комфортной дистанцией для легковых автомобилей можно считать расстояние, которое проедет автомобиль за время не менее 2 секунд. (1.6 + 0.4 = 2).
Время реакции водителя может колебаться от 0,4 до 1,6 секунд, время срабатывания тормозной системы с гидравлическим приводом может составлять от 0,1 до 0,4 секунд. То есть комфортной дистанцией для легковых автомобилей можно считать расстояние, которое проедет автомобиль за время не менее 2 секунд. (1.6 + 0.4 = 2).
В какую сторону смещается прицеп автопоезда на повороте?
| 1. | Не смещается. | |
| 2. | Смещается от центра поворота. | |
| 3. | Смещается к центру поворота. |
При одновременном прохождении поворота с автопоездом вам следует позаботиться об увеличении бокового интервала до этого ТС, так как задние колеса автопоезда (прицепа) смещаются к центру поворота.
При движении в плотном потоке Вы заметили сзади транспортное средство, движущееся на слишком малой дистанции. Как следует поступить, чтобы обеспечить безопасность движения?
1. | Скорректировать скорость движения, ослабив нажатие на педаль газа, чтобы увеличить дистанцию до движущегося впереди транспортного средства. | |
| 2. | Увеличить скорость движения, уменьшив дистанцию до движущегося впереди транспортного средства. | |
| 3. | Предупредить следующего сзади водителя резким кратковременным торможением. | |
| 4. | Допускается любое из перечисленных действий. |
Если при движении в плотном потоке Вы заметили транспортное средство, движущееся сзади на слишком малой дистанции, то целесообразно ослабить нажатие на педаль газа, увеличив тем самым дистанцию до движущегося впереди автомобиля. Таким образом Вы создадите себе резерв времени для применения менее интенсивного торможения при возникновении опасности. Резкое торможение может привести к наезду сзади, а увеличение скорости приведет к уменьшению дистанции спереди.
По какой траектории двигается прицеп легкового автомобиля при прохождении поворота?
1. | Дальше от центра поворота, чем траектория движения автомобиля. | |
| 2. | По траектории движения автомобиля. | |
| 3. | Ближе к центру поворота, чем траектория движения автомобиля. |
При прохождении поворота легкового автомобиля с прицепом вам следует позаботиться об увеличении бокового интервала, так как задние колеса прицепа смещаются к центру поворота.
Количество полос | Боковой интервал
Вопрос: Как выбрать в зимнее время количество полос и боковой интервал между автомобилями? Если при попутном движении не помещается 2 грузовых ТС, или, например, большие автобусы, то следует ли считать, что в попутном направлении дорога имеет одну полосу для движения?
Ответ. Вопрос остается актуальным не только во время зимы, когда часть дороги оказывается «выключенной» из проезжей части по причине снежных заносов, но и для дорог, на которых разметку не видно (стерлась или ее там никогда не было).
Количество полос движения
На вопрос о количестве полос на проезжей части дает ответ пункт 9.1 ПДД:
«Количество полос движения для безрельсовых транспортных средств определяется разметкой и (или) знаками 5.15.1, 5.15.2, 5.15.7, 5.15.8, а если их нет, то самими водителями с учетом ширины проезжей части, габаритов транспортных средств и необходимых интервалов между ними. При этом стороной, предназначенной для встречного движения на дорогах с двусторонним движением без разделительной полосы, считается половина ширины проезжей части, расположенная слева …».
Когда на дороге отсутствует разметка и соответствующие знаки, для чего нужно иметь представление о количестве полос?
Во-первых, это нужно чтобы не выехать на встречную полосу, когда это запрещено (на дорогах с двусторонним движением в четыре и более полосы пункт 9.2 ПДД прямо запрещает выезжать на встречную полосу, даже в том случае, если разметка на дороге отсутствует).
Далее, перед поворотами направо, налево (разворотом) Правила требуют занять соответствующее крайнее положение на проезжей части (крайние правое или крайнее левое, пункт 8.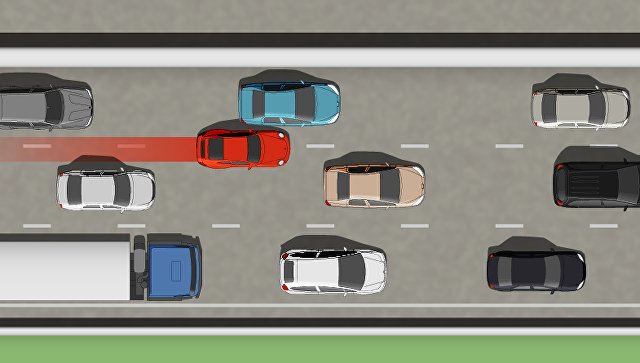 5 ПДД), т.е. перестроиться в нужную сторону.
5 ПДД), т.е. перестроиться в нужную сторону.
Процесс перестроения должен происходить в соответствии с пунктом 8.4 ПДД – уступает тот, кто перестраивается. Подробнее об этом – в статье Одновременное перестроение. Теория и практика.
Таким образом, т.е. во-вторых, количество полос на дороге необходимо знать для прояснения вопросов приоритета во время перестроения и для безопасного выполнения этого маневра.
Каким путем для себя выяснить, где заканчивается попутная половина проезжей части? Для этого мысленно делим вдоль проезжую часть пополам, и «отсекаем» ее левую половину – это будет встречная половина проезжей части.
Далее, берем ширину своего автомобиля и соотносим ее к ширине «своей» попутной половины проезжей части. Прикидываете, сколько ваших машин (или примерно таких) по ширине войдет в это пространство, плюс небольшой запас на безопасный боковой интервал, примерно полметра. Все эти расчеты достаточно условны, не следует пытаться выяснять какие-то точные цифры или разграничения.
Когда дорога не размечена (или под снегом) и нет знаков, определяющих движение по полосам, условную ширину полосы для движения целесообразно брать исходя из ширины вашего (т.е. своего) транспортного средства, но с учетом габаритных размеров соседних машин, плюс нужно учесть боковой интервал (об этом ниже). Для чего это нужно?
«Соседей» с дороги никуда не уберешь, они, ведь, тоже участники движения. И среди них может быть крупногабаритный транспорт, например, автобус, троллейбус, большой грузовик. Вы же, в свою очередь, управляете своим! автомобилем, и траекторию движения выбираете исходя из габаритов ВАШЕГО транспортного средства с учетом окружающей обстановки в данный момент времени.
Там, где не помещаются рядом два автобуса, могут поместиться автобус и легковой автомобиль. Или могут свободно разместиться два легковых автомобиля. И соответственно, там, где для автобуса проезжая часть состоит из одной полосы, для легковушек этих полос уже будет две. Зимой, на заснеженной дороге, рядом этим двум легковым машинам уже может стать «тесно», и для них летние две полосы «превращаются» в одну зимнюю.
После снегопадов, когда дороги должным образом не очищены, четыре реальные полосы движения фактически превращаются в две! И мы видим, как два попутных ряда сливаются в один, машины движутся в одной колее. Но даже эти обстоятельства не отменяют пункт 8.5 ПДД (требование занять крайнее положение перед поворотом направо, налево или разворотом).
Боковой интервал
Теперь о выборе бокового интервала. Что такое боковой интервал, и какой величины должен быть боковой интервал? Боковой интервал при рядном движении – это расстояние между бортами транспортных средств. Транспортные средства, в свою очередь, могут двигаться как попутно, так и навстречу друг другу, и в любом случае боковой интервал – это расстояние между их бортами.
ПДД о величине бокового интервала упоминают лишь условно:
Пункт 9.10: «Водитель должен соблюдать такую дистанцию до движущегося впереди транспортного средства, которая позволила бы избежать столкновения, а также необходимый боковой интервал, обеспечивающий безопасность движения».
Этот необходимый боковой интервал и должен обеспечить безопасное «соседство» смежных транспортных средств, исключить их боковое столкновение. Условно принято считать, что между попутно движущимися с равной скоростью автомобилями, безопасный боковой интервал должен быть не менее 0,5 метра.
Зависит ли боковой интервал от скорости движения и от ширины автомобиля? Да, зависимость прямая. Чем выше скорость и чем автомобиль в габаритах шире, тем боковой интервал должен быть больше. Несоблюдение безопасного бокового интервала чревато боковым столкновением (еще говорят, машины «притерлись» боками).
В каких еще случаях следует увеличивать боковой интервал? Вообще, каких либо определенных ситуаций для этого не предусмотрено, но боковой интервал целесообразно увеличивать на изгибах дороги, во время опережения и обгона (особенно длинномеров), на встречном разъезде, на параллельных поворотах, при движении по кольцу (не стоит «прижиматься»).
Также целесообразно увеличить боковой интервал на скользкой, заснеженной либо покрытой наледью дороге.
Возвращаясь к вопросу читателя в начале статьи о количестве полос движения и выборе бокового интервала, предположим, ваш автомобиль догоняет автобус или троллейбус по заснеженной дороге (проезжая часть ограничена снежными заносами), и есть необходимость его объехать (опередить).
Смотрите на ширину своей половины проезжей части. Если есть возможность опередить слева, не выезжая на встречную половину дороги (т.е. не обгон), значит, смещаетесь левее и опережаете. Если такой возможности нет (узкая дорога), а объехать едущий автобус необходимо, тогда нужно быть уверенным, что выезд на встречную полосу на этом участке дороги не запрещен, и действуете по ситуации.
Соблюдение ПДД – это необходимость. А несоблюдение безопасности чревато столкновением за рамками ПДД. По ситуации – это значит принять меры предосторожности, ведь, скользкая дорога значительно увеличивает остановочный путь, а во время переезда снежных межполосных уплотнений во время перестроения автомобиль может занести, и как следствие – потеря управления.
Какого-либо общего алгоритма действий в таких ситуациях не существует – действовать нужно исходя из условий конкретной дорожной обстановки.
Еще нужно понять следующее. Когда дорогу в четыре полосы (на них выезд на встречную полосу запрещен пунктом 9.2 ПДД) замело снегом, и она фактически стала двухполосной (один ряд – ваш, второй ряд — навстречу), то эта дорога НЕ перестала быть четырехполосной. Она по-прежнему таковой является, но в этой ситуации крайние правые полосы могут быть непригодны для движения (под снегом) только в данный момент времени, пока не очистят.
Более того, на скользкой дороге или во время снежных заносов выезжать на встречную полосу иногда бывает очень нежелательно. Поэтому, в каких-то «снежных» ситуациях движение по одной колее след-вслед может быть единственно целесообразным.
Будьте внимательны за рулем.
Всегда ли в ДТП виноват тот, кто бьет сзади — Российская газета
Автомобиль въехал другому автомобилю в корму. Кто виноват? Ответ не столь однозначен. Разберемся в тонкостях вопроса.
Разберемся в тонкостях вопроса.
Многие автомобилисты ошибочно считают, что при авариях, когда автомобиль ударяет ехавшего впереди соседа по полосе, гарантированно виноват «задний».
Действительно, в пункте 9.10 правил дорожного движения, который не менялся с момента принятия в 1993 году, четко прописано — водитель должен соблюдать такую дистанцию до движущегося впереди транспортного средства, которая позволила бы избежать столкновения, а также необходимый боковой интервал, обеспечивающий безопасность движения.
Тем не менее, как это ни парадоксально, в подавляющем числе случаев при такого рода авариях виновником нередко признают ехавшего впереди. Почему так происходит? Все дело в большом числе оговорок и подпунктов, которые позволяют трактовать пункт правил 9.10 в пользу того, кто, простите за жаргонизм, «догнал» впереди едущего.
К примеру, пункт 9.10 перестает работать в пользу водителя, двигавшегося впереди, когда тот предпринимает экстренное торможение, не требующееся для предотвращения дорожно-транспортного происшествия.
Вот вам типичный пример — водитель резко бьет по тормозам перед перекрестком, подъезжая к нему, скажем, на мигающий «желтый». В случае возникновения ДТП виновником с большой степенью вероятности будет признан именно затормозивший перед светофором, поскольку своими нелогичными и опасными действиями он создал аварийную ситуацию. Впрочем, случается, что при такого рода ДТП виновниками могут признать обоих водителей (например, если тот, кто ехал сзади, не соблюдал дистанцию и скоростной режим).
Еще с большей вероятностью виновниками сочтут так называемых «учителей» — намеренно тормозящих «в пол» перед другими автомобилями, чтобы проучить их водителей за какие-то, как они считают, огрехи. Таким образом, знайте, если машина совершила перед вами внезапную остановку в ситуации, которая этого не требовала, правда на вашей стороне. Останется озаботиться в этом случае лишь доказательной базой — записью с видеорегистратора, близлежащих камер видеонаблюдения или поиском свидетелей — соседей по потоку или прохожих.
Теперь представим себе следующий сценарий. Вы двигаетесь по главной дороге, а другое транспортное средство резко выскакивает перед вами, съезжая с второстепенной дороги. Вы предпринимаете экстренное торможение, но «догоняете» вклинившееся перед вами авто.
Если будут собраны соответствующие доказательства, виновником практически гарантированно признают водителя, бесцеремонно выскочившего прямо у вас по курсу, что предусмотрено положением 8.3 правил дорожного движения.
Согласно ему, при выезде на дорогу с прилегающей территории водитель должен уступить дорогу транспортным средствам и пешеходам, движущимся по ней, а при съезде с дороги — пешеходам и велосипедистам, путь движения которых он пересекает. Если машина перед вами неожиданно перестроилась из другого ряда, резко затормозила, и вы въехали ей в корму, то у вас опять-таки имеются шансы оказаться в этой ситуации правым. Здесь опять-таки желательно иметь видеорегистратор, либо искать камеры наружного наблюдения и свидетелей.
Рассмотрим также и часто встречающуюся ситуацию с так называемыми множественными столкновениями, когда сразу несколько машин въезжают друг другу в корму по цепочке (так называемый «паровозик»).
Практика показывает, что виновником из-за несоблюдения дистанции (пункт 9.10 ПДД) и неспособности снизить скорость при возникновении опасности (пункт 10.1 ПДД) будут признаны хозяева второй, третьей и последующих машин в цепочке. Впрочем, нередко случается и такое. Первая машина въезжает в «зад» соседа по потоку, а тот, следуя по инерции, «догоняет» впереди идущий авто. Невиновность того, кого «толкнули», можно доказать, если провести трасологическую экспертизу или с опорой на показания видеорегистратора и свидетелей. Однако фактически очень часто все участники ДТП получают «обоюдку», когда виновными признают всех участников аварии.
И уже совершенно точно, водителя впереди идущей машины признают виновным, если он совершает наезд на другие машины во время движения задним ходом. Чаще всего такие ДТП возникают при выезде с парковочного места или движении задним ходом на трассе.
Виновником сдававшего назад делает пункт 8.12 ПДД, согласно которому движение транспортного средства задним ходом разрешается лишь при условии, что этот маневр будет безопасен и не создаст помех другим участникам движения. В свою очередь пункт 8.11 оговаривает, что движение задним ходом запрещается на перекрестках и в местах, где запрещен разворот. В любом случае, даже в такой ситуации для доказательства своей правоты понадобится видеорегистратор или показания свидетелей.
|
| ||
|
| ||
|
| ||
|
| ||
|
| ||
|
| ||
|
| ||
|
| ||
|
| ||
|
| ||
|
| ||
|
| ||
|
| ||
| |||
|
| ||
|
| ||
|
| ||
|
| ||
|
| ||
|
| ||
|
| ||
|
| ||
|
| ||
|
| ||
|
|
 07.2021
07.2021
 07.2021
07.2021 02.2015 № 500
02.2015 № 500



 Хватовка. Разработка данного карьера обусловлена строительством автомагистрали «М-12 «Строящаяся Скоростная автомобильная дорога Москва-Нижний Новгород-Казань».
Хватовка. Разработка данного карьера обусловлена строительством автомагистрали «М-12 «Строящаяся Скоростная автомобильная дорога Москва-Нижний Новгород-Казань». местами по Нижегородской области и г. Нижнему Новгороду сохранятся грозы, ожидаются ливни, сильные дожди и град.
местами по Нижегородской области и г. Нижнему Новгороду сохранятся грозы, ожидаются ливни, сильные дожди и град. , проводится «Единый консультационный день» с заместителем начальника Межмуниципального отдела по Арзамасскому и Вадскому районам Управления Федеральной службы государственной регистрации, кадастра и картографии по Нижегородской области Груниным Андреем Михайловичем по вопросам государственной регистрации прав на недвижимость.
, проводится «Единый консультационный день» с заместителем начальника Межмуниципального отдела по Арзамасскому и Вадскому районам Управления Федеральной службы государственной регистрации, кадастра и картографии по Нижегородской области Груниным Андреем Михайловичем по вопросам государственной регистрации прав на недвижимость. 1.3597-20 «Профилактика новой коронавирусной инфекции (COVID-19)» будет организован личный прием граждан заместителем Губернатора Нижегородской области Красновым Дмитрием Германовичем, координирующим работу органов исполнительной власти Нижегородской области
1.3597-20 «Профилактика новой коронавирусной инфекции (COVID-19)» будет организован личный прием граждан заместителем Губернатора Нижегородской области Красновым Дмитрием Германовичем, координирующим работу органов исполнительной власти Нижегородской области 06.2021
06.2021 ГБУ НО «Госветуправление Арзамасского района» проводятся противоэпизоотические мероприятия.
ГБУ НО «Госветуправление Арзамасского района» проводятся противоэпизоотические мероприятия.

Какая дистанция должна быть между автомобилями: безопасное расстояние при движении
29.03.2021
Дорожное движение имеет большое количество нюансов. И водителю всегда необходимо контролировать обстановку на дороге, чтобы не допустить аварийных ситуаций. Любую проблему проще предупредить, поэтому при выезде на трассу или на улицы города, нужно быть максимально внимательным и осторожным, а любые маневры применять только тогда, когда это целесообразно.
У начинающих водителей часто возникает вопрос: какая дистанция должна быть между автомобилями. И это действительно важно, поскольку из-за несоблюдения интервалов чаще всего происходят дорожно-транспортные происшествия. Одним из первых нюансов, которым учат инструкторы в автошколах, является соблюдение дистанции. Кроме того, необходимо учитывать и боковой интервал. Какой же должна быть дистанция между авто в различных ситуациях?
Что такое дистанция и боковой интервал?
Дистанция в дорожном движении – это расстояние между транспортными средствами на полосе, которое позволяет безопасно двигаться по дороге. Необходимость соблюдать дистанцию критически важна для водителей, поскольку чаще всего на дорогах происходят именно столкновения. Поэтому вам обязательно нужно определять безопасную дистанцию и соблюдать ее, чтобы не создавать аварийных ситуаций.
Боковой интервал между автомобилями – это боковое расстояние между машинами. Особенно важно соблюдать его при перестроениях. Нередко начинающие водители не соблюдают условия безопасного перестроения и не контролируют наличие других авто в зеркалах. Замечают их только тогда, когда остановиться уже невозможно. Поэтому необходимо быть бдительными, если вы решили менять полосу движения.
Какими должны быть дистанция и боковой интервал?
Касаемо соблюдения дистанции, ПДД гласит, что водители должны интуитивно определять безопасную дистанцию, в зависимости от дорожных условий. Безопасной дистанцией считается расстояние между авто, при котором тормозного пути автомобиля будет достаточно для полной остановки транспортного средства до контакта, при условии технически исправных тормозов.
Итак, какое расстояние должно быть между машинами? Во-первых, стоит учесть, что безопасная дистанция может быть разной, в зависимости от погодных условий и места, где вы едете. Рекомендуется рассчитывать дистанцию, как скорость разделенная на 2. Например, при езде по трассе при условии сухого дорожного покрытия, при скорости 60 км/ч, безопасная дистанция составит 30 м. Аналогичную дистанцию рекомендуется соблюдать и в населенных пунктах, где трафик не перегруженный.
Если же на дороге большое количество автомобилей и образуется пробка, вы будете ехать с низкой скоростью. Однако здесь всегда есть риск, что кто-то попытается перестроиться на вашу полосу. в таком случае рекомендуется придерживаться дистанции 3-4 метра.
Также есть случаи, при которых дистанцию рекомендуется увеличить:
- При плохой видимости – в случае тумана, дождя, снегопада;
- На мокром дорожном покрытии;
- Если приходится ехать в веренице из 5 и более авто.
Если водитель впереди увеличил скорость, не стремитесь его догонять. Если он увеличил дистанцию, продолжайте соблюдать свой скоростной режим.
В случае с боковым интервалом, принцип аналогичный. Его также интуитивно определяет водитель. Рекомендуется ехать с интервалом, равным 1/2 ширины автомобиля. Например, если ширина авто – 1.8 м, лучше придерживаться расстояния 0.9 м. Штраф за несоблюдение бокового интервала не предусмотрен.
Рейтинг: 5/5 — 3 голосов
Другие страницы
lateral% 20spacing — определение английского языка, грамматика, произношение, синонимы и примеры
Гробница была разрушена во время Французской революции, но позже, отремонтированы и выставлены сегодня.
WikiMatrix
Это позволит нам — с оговорками, я представлю позже , чтобы завершить разбирательство в Судебной камере в
MultiUn
Первоначально награды были сосредоточены на компьютерных играх, но затем были расширены, чтобы включить также и консольные игры, благодаря успеху таких игровых консолей, как Sega Master System и Sega Mega Drive в Соединенном Королевстве.
WikiMatrix
JMA также модернизировал Dolphin до тайфуна через несколько часов позже .
WikiMatrix
Наблюдения должны поступить в Комиссию не позже , чем через 10 дней после даты публикации.
ЕврЛекс-2
Получателям будет предложено принять свои знаки отличия на церемонии, которая состоится в позже числа. Впервые присужденный в 1967 году, в год столетия Канады, Орден Канады положил начало созданию собственной системы наград нашей страны.
Гига-френ
Таким образом, Иисус и его апостолы учили, что он «Сын Божий», но позже церковных деятеля развили идею «Бога-Сына».
jw2019
Позже, , он играл за Дебрецену Дожу, прежде чем перейти в KC Veszprém, где он достиг своих лучших результатов в своей карьере, выиграв пять чемпионатов Венгрии и пять титулов в Кубке Венгрии.
WikiMatrix
Чанг, делегат из Китая, позже заявил, что «цель [положения о моральных правах] заключалась не только в защите художников, но и в защите интересов каждого.
MultiUn
Воцарилась тишина, за ней последовал момент спустя калишит, который издал отчаянный стон, когда его глаза закатились, и он безвольно ударился о выжженный газон.
ханглиш
Они, в частности, подчеркнули, что некоторые организации взяли за правило выпускать свои пресс-релизы сначала на английском языке, а перевод на французский будет через несколько дней позже , или, что еще хуже, такие выпуски выпускаются только на английском
MultiUn
Однако три других в настоящее время проходят оценку Федеральной канцелярии и должны стать ценными инструментами позже года.
ЕврЛекс-2
Он стал единственным певцом в группе, хотя Фариан , позже показал, что Бобби почти не участвовал в вокальных записях группы, а сам Фариан исполнял мужские партии в песнях в студии.
WikiMatrix
Как отмечал Ленин позже : «Либо вши победят социализм, либо социализм победит вшей».
WikiMatrix
Оглядываясь назад, можно сказать, что важность дизайна такси во многом объяснялась тем фактом, что он включал в себя поперечно установленный двигатель / трансмиссию, применяя базовую архитектуру, которая была бы воспринята как революционная, когда Алек Иссигонис применил ее в BMC Mini пятьдесят лет спустя . .
WikiMatrix
Сталин, его первый действующий президент, использовал принципы демократического централизма, чтобы превратить свой пост в должность лидера партии, а затем позже лидера Советского Союза.
WikiMatrix
Полет ранее был приостановлен во время захода на посадку к проливу Мак-Мердо, чтобы выполнить снижение с помощью маневра в форме восьмерки через разрыв в нижней границе облаков (, позже , по оценкам, примерно на высоте от 2 000 до 3 000 футов (от 610 до 910 м)). )) во время полета, чтобы установить визуальный контакт с наземными ориентирами и обеспечить лучший обзор для пассажиров.
WikiMatrix
Дикинсон и другие критики осудили использование мелодии позже в патриотическом гимне «Я клянусь Тебе, страна моя» — несмотря на полное соучастие Холста.
WikiMatrix
Уполномочивает Правление Объединенного пенсионного фонда персонала Организации Объединенных Наций повысить обычный возраст выхода на пенсию до 65 лет для новых участников Фонда, причем не на позже , чем с 1 января 2014 года, если Генеральная Ассамблея не примет решение о соответствующем повышении возраста обязательного выхода на пенсию. разделения;
UN-2
Позже, вечером, я выступал с той же речью в Ратуше для европейцев, 35 из которых присутствовали.
jw2019
Позже , как сообщается, она была продана разборке судов в Алиаге, Турция, и переименована в Antic.
WikiMatrix
Предлагается проконсультироваться с ИТЦ или его вспомогательными органами, чтобы определить, сочтут ли государства-члены полезной разработку одного или нескольких многосторонних соглашений на основе Рекомендации L и сделать ссылку на транспортные нормы и стандарты, разработанные ЕЭК ООН, которые в настоящее время не имеют обязательной силы (рекомендации
MultiUn
Части плана полета самолета, соответствующие маловысотным боковым траекториям , могут содержать проходы, имеющие боковую свободу , которая ограничена рисками столкновения с землей или препятствиями на земле.
патенты-wipo
Я имею в виду, что рано или поздно я должен или позже .
OpenSubtitles2018.v3
С хронологической точки зрения, одна станция (Кодиат Абд Эль Хак) демонстрирует бесспорный перетягивание, маленькую антилопу с ослабленными ногами перебивает большой слон, хотя другая, кажется, была на позже на , чем она.
WikiMatrix
Письмо об интерпретации 4-358 (I) — Ресурсы
Версия в формате PDF, 65 КБ
Для просмотра PDF на этой странице вам понадобится Adobe Acrobat Reader.
Министерство транспорта США
Федеральное управление шоссейных дорог
1200 Нью-Джерси-авеню, SE.
Вашингтон, округ Колумбия 20590
5 марта 2009 г.
В ответ обратитесь к: HOTO-1
Роберт Э. Вилак младший, P.E.
Инженер-транспортник
Департамент транспорта округа Марион,
412 S.E. 25 -й проспект
Окала, Флорида 34471
Уважаемый г-н.Вилак:
Благодарим вас за ваше письмо от 24 февраля с просьбой дать официальную интерпретацию пункта F второго Стандартного положения в Разделе 4D.15 Руководства по унифицированным устройствам управления движением (MUTCD), в котором говорится:
F. Требуемые сигнальные поверхности для сквозного движения на любом одном заходе на посадку должны располагаться на расстоянии не менее 2,4 м (8 футов) друг от друга при измерении по горизонтали перпендикулярно подходу между центрами сигнальных поверхностей.
Ключевая фраза в этом предложении — «требуемые сигнальные лица».«В то время как некоторые дополнительные сигнальные грани являются полностью необязательными, пункт D.1.b. того же Стандарта требует наличия дополнительной сигнальной грани на ближней стороне, когда оба из двух основных сигнальных лиц для основного движения находятся на расстоянии более 180 футов от упора. Вы спрашиваете, должно ли минимальное боковое расстояние в 8 футов между «необходимыми сигнальными гранями для сквозного движения» применяться одновременно к первичным поверхностям на дальней стороне и «требуемым» дополнительным поверхностям на ближней стороне.
Назначение пункта F — обеспечить адекватное боковое разделение между непосредственно примыкающими сигнальными гранями для движения.Пункт F не является требованием, чтобы одна дополнительная поверхность на ближней стороне располагалась непосредственно на одной линии в продольном направлении с одной из основных поверхностей на дальней стороне. Дополнительная боковая поверхность, если она предусмотрена из-за проблемы с продольным расстоянием согласно пункту D.1.b. или по любой другой причине, может располагаться в любом боковом положении при условии соблюдения инструкций далее в Разделе 4D.15:
Если используются дополнительные сигнальные лица, их следует располагать так, чтобы обеспечить оптимальную видимость для контролируемого движения.
Однако, если для сквозного движения предусмотрены две или более дополнительных боковых сигнальных поверхности, то эти несколько боковых сторон должны быть расположены сбоку на расстоянии не менее 8 футов друг от друга.
Надеюсь, что эта интерпретация ответит на ваш вопрос. Спасибо, что написали на эту тему. Если у вас есть какие-либо вопросы, свяжитесь с г-ном Скоттом Уэйнрайтом из нашего персонала по электронной почте [email protected] или по телефону 202 366 0857.Обратите внимание, что мы присвоили вашему запросу следующий официальный номер интерпретации и заголовок: «4-358 (I) — Боковое расстояние между гранями дополнительных сигналов». Пожалуйста, обращайтесь к этому номеру в любой будущей переписке по этому вопросу.
С уважением,
Оригинал подписан:
Пол Пизано
И.о. директора Управления транспортных операций
Входящее письмо
Транспортный департамент округа Мэрион
412 с.E. 25 th Avenue
Окала, Флорида 34471
(352) 671-8686
Факс (352) 671-8687
Одна команда… Одна миссия
24 февраля 2009 г.
Управление транспортных операций (HOTO)
Федеральное управление шоссейных дорог, остановка почты: E84-402
1200 Нью-Джерси-авеню, S.E.
Вашингтон, округ Колумбия 20590
RE: Запрос на толкование — MUTCD Раздел 4D.15
Мистер Уэйнрайт:
Обращаюсь к вам с просьбой разъяснить этот раздел, связанный с использованием и расположением «дополнительных» сигнальных головок.MUTCD сообщает следующее:
D. За исключением случаев, когда ширина пересекающейся проезжей части или другие условия делают это физически непрактичным:
- Сигнальная панель, установленная в соответствии с требованиями к указателям левого поворота (см. Раздел 4D.06) и указателям правого поворота (см. Раздел 4D.07), и по крайней мере одна, а лучше обе из двух требуемых сигнальных поверхностей для основного движения на подходе должны располагаться:
- Не менее 12 м (40 футов) за стоп-линией.
- Не более 55 м (180 футов) за стоп-линией, если не предусмотрена дополнительная боковая сигнальная поверхность.
- Как можно ближе к линии обычного обзора водителя, если он установлен над проезжей частью.
- Если ближайшая сигнальная поверхность расположена на расстоянии от 45 до 55 м (от 150 до 180 футов) за стоп-линией, инженерная оценка условий, включая наихудшие условия видимости, должна использоваться для определения того, предусмотрена ли дополнительная ближняя граница. Боковое сигнальное лицо было бы полезно.
F. Требуемые сигнальные поверхности для сквозного движения на любом одном заходе на посадку должны располагаться на расстоянии не менее 2,4 м (8 футов) друг от друга при измерении по горизонтали перпендикулярно подходу между центрами сигнальных поверхностей.
Мы столкнулись с несколькими ситуациями, когда превышается максимальное расстояние 180 футов за стопорной планкой, а конструкция и установка предусматривают дополнительную сигнальную головку над стопорной планкой, которая находится непосредственно на одной линии с дальней сигнальной головкой.
Можно ли в этом приложении упустить требуемый интервал 8 футов между центрами сигнальных граней или дополнительные ближние сигнальные головки должны быть установлены слева или справа от дальних боковых сигналов?
Спасибо за помощь в этом вопросе,
С уважением,
Роберт Э.Вилак, младший, П.
Инженер-транспортник
тел: (352) 671-8686
факс: (352) 671-8687
электронная почта: [email protected]
«Влияние бокового зазора на начало пружины» Кайл Луи Гроссарт
Абстрактные
Спринтеры всегда стремятся улучшить свое время, от выстрела до финиша. Эффективный старт может привести к более быстрому достижению максимальной скорости и сокращению времени финиша.Новые разработки в области стартовых колодок, в частности, ширины педали стартового блока, позволили варьировать положение ступней в блоках. В этом исследовании была предпринята попытка найти оптимальное расстояние для спринтеров уровня колледжа. В исследовании приняли участие тринадцать мужчин-спринтеров из колледжа (средний возраст = 23,08 года). Испытуемые самостоятельно выбирали расстояние между блоками в продольном направлении с 3 различными боковыми позициями для тестирования.В положении 1 ступни были размещены настолько узкими, насколько позволял стартовый блок, имитируя ширину традиционного набора блоков. Положение 2 определялось шириной бедер человека, расстоянием между правым ASIS и левым ASIS. Положение 3 было предпочтительной шириной стопы испытуемого, определяемой при выполнении вертикального прыжка. Были проведены измерения максимальной силы на блоки в момент старта, а также время до 10 м. Ни пиковая сила, ни время до 10 м не различались в зависимости от условий (p =.887, р = 0,135). Нормальное состояние, положение 1 (20 см), было измерено как самая узкая ширина, а положение 3 (среднее значение = 37,6 см) было самым широким у всех испытуемых. Использование более широких педалей на стартовых блоках — довольно новая идея в легкой атлетике, и это то, с чем многие спортсмены не имели возможности тренироваться. Поскольку старый стиль стартовых блоков допускал только узкую стойку, это то, к чему легкоатлеты привыкли, и, возможно, может объяснить, почему не было заметной разницы между тремя стартовыми позициями.После периода адаптации спортсменов к новым стартовым блокам следует провести дополнительные исследования.
Колледж и факультет
Науки о жизни; Физические упражнения
Права
http://lib.byu.edu/about/copyright/
Ученые BYU Архивная ссылка
Гроссарт, Кайл Луис, «Влияние бокового зазора на начало пружины» (2010). диссертаций и диссертаций . 2554.
https://scholarsarchive.byu.edu/etd/2554
Дата отправки
08.07.2010
Ручка
http: // hdl.lib.byu.edu/1877/etd3750
Ключевые слова
стартовых блока, пиковая сила, время до 10 м, боковая ширина стопы
Реакция арахиса на севооборот, расстояние между капельными трубками и скорость орошения при глубоком подповерхностном капельном орошении
Долгосрочная урожайность при различных севооборотах с подпочвенным капельным орошением (SSDI) не известна для юго-востока США. Система SSDI была установлена в 1998 году на суглинистой песчаной почве Тифтон с пятью севооборотами, двумя боковыми интервалами капельных трубок и тремя уровнями орошения.Севооборот варьировался от непрерывного арахиса ( Arachis hypogaea л) до четырех лет между арахисом. Боковые стенки были установлены под каждым рядком (0,91 м) и попеременными междурядьями (1,83 м). Посевы орошались ежедневно из расчета 100, 75 и 50% расчетного водопотребления сельскохозяйственных культур. Боковые стволы с шагом 1,83 м дали такой же урожай, как отводы с шагом 0,91 м за девять из десяти лет. Обработка 50, 75 и 100% орошения составила в среднем 3263, 3468 и 3497 кг / га соответственно. Не было разницы в урожайности между 75 и 100% обработкой орошения, что подразумевает экономию воды на 25%.Севооборот повлиял на урожайность арахиса в семи из восьми лет, а непрерывный арахис дал самый низкий урожай за все годы. По мере того, как время между посевами арахиса увеличивается, урожайность арахиса увеличивается. Обработка орошением не повлияла на общие здоровые зрелые ядра (TSMK). Поперечный интервал влияет на TSMK в 20% случаев, а севооборот влияет на TSMK в 90% случаев. Непрерывный севооборот арахиса показал самый низкий TSMK, при этом более высокий TSMK наблюдался по мере увеличения времени между посевами арахиса. Не было доказательств того, что какой-либо один севооборот отрицательно повлиял на гранулометрический состав зерна, за исключением арахиса непрерывного действия.При использовании SSDI можно сэкономить 25% воды для орошения, установить отводы в чередующихся междурядьях и чередовать с арахисом каждые три года без отрицательного влияния на урожайность или сорт арахиса.
Производство арахиса покрывает чуть более 322 000 га в трех штатах Алабама, Флорида и Джорджия, и только 28% этих акров орошаются (USDA, 2009). Только 10% от общего количества орошаемых гектаров в Джорджии орошались капельным, капельным или микро-дождеванием, в то время как во Флориде более 220 000 га использовали тот или иной тип капельного или капельного орошения (USDA, 2009).Из-за дороговизны установки капельной системы предполагается, что большая часть этих капельных систем предназначена для выращивания ценных овощных культур. Неизвестно, можно ли и сколько из этих капельных систем можно использовать для выращивания арахиса или других традиционных пропашных культур, таких как хлопок ( Gossypium hirsutum L.) или кукуруза ( Zea mays L.).
Экономическое моделирование показало, что подземное капельное орошение (SSDI) будет более прибыльным для небольших территорий (<30 га) из-за меньших затрат на единицу площади земли и меньших затрат на перекачку по сравнению с фиксированными или буксируемыми системами с центральным шарниром.Как подчеркивают Bosch и др. . (1998) и О’Брайен и др. . (1998), системы SSDI имеют почти статичную стоимость гектара по сравнению с верхними дождевальными системами (центральные шарниры), где стоимость гектара уменьшается по мере увеличения длины системы с охватом большей площади. Верхние дождевальные оросительные системы являются наиболее распространенными в районе трех штатов, потому что они просты в сборке, долговечны, не требуют сложных систем фильтрации и знакомы с эксплуатацией и обслуживанием.Одна из основных проблем, связанных с верхними дождевальными системами, заключается в том, что как только вода выходит из верхней дождевальной системы, ее судьба может зависеть от условий окружающей среды, так что вода может не попасть в заданную цель, но будет потеряна из-за ветра и испарения, прежде чем она достигнет поверхности почвы и станет доступны для использования в сельскохозяйственных культурах. Таким образом, подземное капельное орошение может обеспечить стабильно высокие урожаи при сохранении почвы, воды и энергии. Некоторые из основных преимуществ капельного орошения включают точное размещение воды и химикатов, низкие затраты на рабочую силу, а также меньший сток и эрозию по сравнению с системой дождевания сверху.Эти системы SSDI могут часто подавать воду в корневую зону, тем самым снижая риск циклического водного стресса, типичного для других систем орошения. Исследования показали, что урожайность и качество урожая могут быть увеличены и что SSDI можно использовать для хлопка (Bucks et al ., 1988; Henggeler, 1988, Nuti et al., 2006, Dougherty et al ., 2009 ) и кукурузы (Mitchell, 1981; Mitchell and Sparks, 1982; Powell and Wright, 1993).
Эти системы SSDI можно адаптировать к различным размерам и формам полей, что делает их важным экономическим фактором, особенно на юго-востоке.Это экономическое преимущество становится еще более очевидным при рассмотрении варианта разработки системы SSDI для эффективного покрытия поля неправильной формы, которое не может быть полностью покрыто системой спринклерного типа (Bosch et al ., 1998). При правильной конструкции SSDI эти системы могут обеспечивать достаточным количеством воды различные поля в зависимости от площади, почв и видов сельскохозяйственных культур.
Отводы капельной трубки установлены на 0.Глубина почвы 2 и 0,3 м (Bucks et al., 1986; Tollefson, 1985; Phene et al ., 1987; Camp et al. ., 1989) на хлопке, кукурузе, фруктах и овощах. Отводы капельного орошения были расположены на расстоянии 1, 2 и 3 м друг от друга, при этом урожайность уменьшалась по мере увеличения поперечного расстояния до более чем 2 м (French et al ., 1985; Lamm et al., 1992; Powell and Wright, 1993 ; Camp et al., 1997; Enciso et al ., 2005). Капельные трубки закапывались или укладывались на поверхность почвы на различных расстояниях по бокам, т.е.е., каждый ряд или чередующиеся борозды при непрерывном чередовании хлопка или хлопка-кукурузы-арахиса (Camp et al ., 1993; Camp et al ., 1997; Dougherty et al ., 2009; Sorensen et al ., 2008). У непрерывного хлопка с чередованием поперечного расстояния между рядами наблюдалась изменчивость из года в год из-за климатических условий, но урожайность орошаемого хлопка была выше, чем урожайность неорошаемого, особенно в засушливые годы (Dougherty et al., 2009). Сравнение альтернативного междурядья с поперечным расстоянием между рядами показало отсутствие разницы в урожайности при непрерывном хлопковом севообороте или севообороте хлопка-кукурузы-арахиса (Camp et al., 1993; Camp et al ., 1997; Соренсен и др., 2008).
В связи с растущей заботой об экономии воды в регионе трех штатов (Алабама, Джорджия и Флорида), использование SSDI из-за большей эффективности орошения этих систем может представлять большой интерес для отдельных производителей, водохозяйственных агентств и агентств по охране окружающей среды. , и агентствами, определяющими политику. Имеется мало долгосрочных данных об урожайности арахиса с SSDI на юго-востоке, чтобы дать рекомендации по управлению.Таким образом, целью этого исследования было определить долгосрочную реакцию урожайности арахиса на: 1) три нормы орошения, 2) два боковых междурядья и 3) пять севооборотов с использованием SSDI за 10-летний период.
Место исследования было расположено в округе Террелл недалеко от Сассера, Джорджия, на супесчаной почве Тифтон (тонкосуглинистые, каолинитовые, термические плинтусы Кандиудультов) с уклоном от 2 до 5%. Система SSDI была установлена в 1998 году на неорошаемых сельскохозяйственных угодьях, которая состояла из трех уровней орошения, пяти севооборотов, двух боковых интервалов капельных трубок и трех повторений для всего 90 отдельных участков.Хлопок был посажен за два года до установки системы SSDI. Право собственности на землю изменилось так, что долгосрочные севообороты были недоступны, а нынешний владелец не выращивал арахис с 1993 года. Прямоугольник площадью 6,8 га был разделен на три равные площади, называемые ярусами. Между ярусами, по бокам и по краям рядов сельскохозяйственных культур для мест разворота оборудования были предусмотрены проезды (минимум 12,2 м). Каждому ярусу SSDI (38 м на 274 м) случайным образом был назначен уровень орошения. Уровень SSDI состоял из трех блоков (репликаций), пяти севооборотов и двух тонкостенных капельных интервалов по бокам, всего 30 участков на ярус.Уровни орошения составляли 100%, 75% и 50% от расчетного водопотребления сельскохозяйственных культур (Sorensen et al. , 2001).
Пять севооборотов включали арахис непрерывного действия (PPP), хлопок-арахис (CP), кукурузу-арахис (MP), хлопок-кукурузу-арахис (CMP) и хлопок-кукурузу-кукурузу-арахис (CMMP) (Таблица 1 ). Все культуры были посеяны с междурядьем 0,91 м в однорядную схему. У двух боковых обработок капельных трубок были капельные трубки, установленные под каждым рядом культур (узкие, 0.91-м) и в междурядьях чередования культур (широкие 1,83 м). На каждом узком ряду подучастка было шесть рядов культур с одной боковой трубкой для капельницы, установленной под каждым рядом, и они были реплицированы три раза на каждом уровне (повторение на блок). На каждом подучастке с широкими рядами было 10 рядов культур с пятью боковыми трубками для капельниц, установленных в чередующихся срединных рядах ряда культур, и повторялись три раза, по одной репликации в каждом блоке. Соренсен и др. . (2001) подробно описывает обработки, критерии проектирования ирригационной системы и контроль орошения.Тонкостенная капельная трубка (Super Typhoon, Netafim Irrigation, Inc., Фресно, Калифорния; www.netafim-usa.com) имела толщину стенки 0,254 мм и эмиттеры, расположенные через каждые 46 см, с расходом 1,5 л / ч. на эмитента. Все тонкостенные капельные трубы были заглублены примерно на 30 см с использованием модифицированной стойки рыхлителя.
Таблица 1.Описание севооборота для исследований в области орошения в Сассере, Джорджия.
Поливная вода применялась ежедневно на основе замены расчетного водопотребления сельскохозяйственных культур на арахис (Таблица 2).Ежедневно регистрировались температура воздуха (максимальная, минимальная и средняя), относительная влажность, общая солнечная радиация и осадки. С 1998 по 2003 год метеорологические данные собирались с использованием модулей программируемого логического управления (ПЛК). Эта система работала, но была уязвима для молний. Весной 2004 года эта система ПЛК была заменена более надежной системой регистрации данных (Campbell Sci., Inc, Logan, UT; CR23X). Суточная потенциальная эвапотранспирация (ET o ) оценивалась с использованием модифицированного уравнения Йенсена-Хейза, скорректированного с учетом местных условий (Jensen and Haise, 1963).Суточные коэффициенты урожая, K c , были определены путем деления расчетной средней суточной арахиса (Stansell et al. ., 1976), хлопка (Harrison and Tyson, 1993) и кукурузы (Lambert et al. , 1988) значения водопользования по суточной оценке архивных данных ET или за тот же период времени. Затем суточное значение ET o было умножено на суточное значение K c , чтобы оценить суточное восполнение воды для каждой культуры (расчетное значение ET a ), которое определяется как 100% -ный уровень орошения.Два других уровня орошения были определены путем умножения 100% уровня орошения на 75 и 50% соответственно. Продолжительность полива для каждой поливной обработки рассчитывалась на основе расчетных суточных значений ET и для внесения желаемой глубины воды. Орошение не применялось, если количество осадков превысило расчетное водопотребление сельскохозяйственных культур в этот день. Максимум 10 мм осадков будет использоваться в качестве «переходящего остатка» для остановки орошения на короткий промежуток времени после выпадения осадков.Ежедневные значения ET a вычитались из «переходящего» значения до тех пор, пока его значение не обнулялось, после чего возобновлялись события полива. 10-миллиметровый «перенос» составляет около 25% истощения почвы для этого типа почвы.
Таблица 2.Сроки посадки и сбора арахиса, количество осадков, орошение и сорт, выбранный по годам для исследования орошения в Сассере, Джорджия.
Известь применялась на всех участках и во все годы, как было определено с помощью почвенного теста, для поддержания pH почвы примерно на уровне 6.5. Подготовка посевного ложа для всех культур состояла из одного-двух проходов (один раз осенью и один раз весной) с использованием экспериментального почвообрабатывающего оборудования (USDA-ARS-Национальная исследовательская лаборатория арахиса), которое в основном позволяло обрабатывать верхние 10-15 см. почвы. Это оборудование преобразовало бы почву в одну грядку шириной около 1,4 м. Это оборудование также обеспечивало возможность контролируемого движения, так что ни одно колесное оборудование не наезжало на заглубленные боковые позиции. Небольшой полевой культиватор использовался для разрушения почвенной корки, внесения гербицидов и борьбы с сорняками перед посевом любых культур.После уборки пожнивные остатки скашивают (хлопок и кукуруза), слегка обрабатывают дисковой бороной, а затем повторно засыпают, как описано ранее.
Единственный используемый сорт арахиса, «Джорджия Грин» (Branch, 1996), был посажен в период с 1 мая st до 12 th (в зависимости от погодных условий) с помощью сеялки вакуумного типа (Monosem, ATI., Inc., Lenexa, KS) примерно 20 семян м -1 на междурядье 0,91 м (Таблица 2).В каждый соответствующий год для лечения применялись одни и те же приложения по борьбе с сорняками, насекомыми и болезнями в соответствии с общими рекомендациями, изложенными на этикетках отдельных продуктов или рекомендациями Службы сельскохозяйственных знаний Университета Джорджии (Prostko, 2004). Сроки сбора урожая основывались на оптимальной зрелости урожая, определяемой методом зачистки корпуса (Williams and Drexler, 1981). Ряды урожая копали 2-рядным инвертором и убирали 2-рядным комбайном. Вес образцов был записан и впоследствии разделен таким образом, что от каждого образца на участке была отобрана подвыборка весом от 4 до 7 кг.Каждая подвыборка была оценена и очищена, чтобы определить сорт фермерского поголовья и гранулометрический состав, соответственно. Выход стручков был основан на общей массе образца, доведенной до 7% влажности. Сорт фермерского поголовья и распределение ядер по размеру были определены с использованием процедур, установленных Министерством сельского хозяйства США (USDA, 1998). Валовая выручка была определена с использованием средней рыночной цены за 2008 год в размере 0,45 доллара США за -1 арахиса фермерского поголовья (Департамент сельского хозяйства штата Джорджия, 2009).
Из-за ограниченной площади земель не каждый севооборот проводился ежегодно.Следовательно, не каждую комбинацию севооборота арахиса можно анализировать путем ротации каждый год. Для анализа данных об урожайности и содержании арахиса (Statistix9, 2008) в отношении нормы орошения (ярусов), севооборота и поперечного расстояния использовался факторный план процедуры общего дисперсионного анализа. Данные по урожайности и сортам были проанализированы по отдельным годам, режиму орошения, севообороту и поперечному расстоянию в пределах и по годам, если применимо. Различия между средними значениями урожайности и качества определяли с использованием множественного сравнения Tukey HSD, когда ANOVA F-тест показал значимость ( P ≤ 0.05).
Меньшее количество осадков было в 2002 году — чуть менее 300 мм за вегетационный период (день года, 121–258 динаров) с 1 мая -го с до середины сентября. Наибольшее количество осадков выпало в сезоне 2005 года. Среднее количество осадков за все десять вегетационных сезонов составило 477 мм. В таблице 2 также показаны объемы орошения, применяемые в течение вегетационного периода для различных лет, и уровни орошения. За 10-летний период 100% орошение составляло в среднем 295 мм орошения за вегетационный период.Обработка 75 и 50% орошения составляла в среднем 213 и 154 мм орошения за вегетационный период для фактической обработки орошения 72 и 52%, соответственно. Эти проценты были очень похожи на рассчитанные обработки, подразумевая, что ирригационная система хорошо работала в течение этого 10-летнего периода. На рисунке 1 показаны кумулятивные осадки, орошение и расчетное суммарное испарение (ЕТа) для года с низким (2002 г.), высоким (2005 г.) и средним (2006 г.) количеством осадков. В 2002 году ожидалось, что потребуется более высокий объем орошения по сравнению с другими годами из-за небольшого количества осадков.На Рисунке 1A орошение было намного меньше расчетного водопотребления, однако совокупное орошение плюс количество осадков близко соответствовало расчетному ET a урожая. На Рисунке 1B показаны кумулятивные ET и , орошение и осадки плюс орошение для 2005 года с наибольшим количеством осадков, а на Рисунке 1C показаны те же параметры для 2006 года, когда количество осадков было несколько средним. На рисунках 1B и 1C показатели орошения и дождевых осадков были намного больше, чем расчетное значение ET на урожая.Количество осадков может быть довольно большим при длительных периодах засухи между дождевыми дождями. Таким образом, время и количество полива в значительной степени зависят от количества, интенсивности и количества дождя.
Рис. 1.
Расчетное кумулятивное ET a и измеренное кумулятивное орошение и орошение плюс количество осадков в течение 2002 (A), 2005 (B) и 2006 (C) лет для низких, высоких и средних осадков между днями года. 121 по 257.
Рис. 1.
Расчетное кумулятивное ET a и измеренное кумулятивное орошение и ирригация плюс количество осадков в течение 2002 (A), 2005 (B) и 2006 (C) лет для низкого, высокого и среднего количества осадков между днями год 121 по 257.
В таблице 3 показаны значения вероятности дисперсионного анализа для урожайности, сортности и распределения ядер по размеру. Было только два года, когда оросительная обработка показала значительную разницу в урожайности, 2003 и 2006 гг.Эти два года не считались годами с малым количеством осадков; Фактически, в 2003 году было зафиксировано второе по величине количество осадков, в то время как 2006 год был близок к среднему (см. Рисунок 1C). Казалось бы, эффекты обработки для орошения будут иметь место в годы засушливых дождевых осадков 2000 и 2002 гг. Как обсуждалось ранее, совокупное орошение в 2002 г. было намного меньше, чем ET a с орошением плюс количество осадков, почти равным расчетному ET a от обрезать. Однако в 2002 году не было снижения урожайности между уровнями орошения, что указывает на то, что количество дождевых осадков плюс ирригация казалось достаточным даже для 50% -ного уровня орошения.Данные по урожайности показывают, что в 2003 и 2006 годах были измерены более низкие урожаи при 50% орошении по сравнению с 75 и 100% орошением (Таблица 4). Поскольку только два года из десяти показывают более низкую урожайность при 50% -ной ирригационной обработке и отсутствие значительной разницы в урожайности между 75-процентной ирригационной обработкой, было бы разумным сделать вывод, что орошение 75% расчетной ЕТа будет эффективным уровнем полива . Орошение при 75% расчетного значения ET — означало бы экономию воды на 25% без снижения урожайности сельскохозяйственных культур.Кроме того, в зависимости от характера выпадения осадков может быть даже возможно сократить орошение на целых 50% от расчетной ЕТа без снижения урожайности для 50% усилий по сбережению воды; однако риск снижения урожайности будет увеличиваться по мере увеличения продолжительности засухи. Характер осадков может объяснить причину реакции урожайности на орошение в одни годы (2003 и 2006 годы), а в другие — нет. Месячные данные об осадках показывают, что в 2006 г. и июнь, и июль были очень засушливыми месяцами с более чем 70 днями подряд, и только один случай выпадения осадков превышал 5 мм.В 2003 году ежемесячные суммарные показатели за вегетационный период превышали 130 мм / месяц, однако было несколько периодов до 17 дней подряд без осадков, за которыми следовали интенсивные ливни. В 2002 году осадков было меньше всего в мае и июне, но июль был влажным. Эти примеры показывают, что совокупное количество осадков за год или месяц не обязательно коррелирует с урожайностью. Следовательно, отдельные случаи выпадения дождя (время), интенсивность и общее количество могут стать проблемой для планирования полива и определения окончательной урожайности.
Таблица 3. Значения вероятностидля урожая, сорта (TSMK, масличный запас) и распределения зерна по размеру (большие, средние и единицы) по отношению к орошению, севообороту и поперечному расстоянию для арахиса с 1999 по 2007 год в Сассере, Джорджия.
Таблица 4.Годовая урожайность арахиса и средняя проектная урожайность для уровня орошения, межосевого расстояния и севооборота по годам с 1999 по 2007 год для Сассера, Джорджия.
За 10-летний период этого исследования, 50-, 75- и 100-процентная оросительная обработка составила в среднем 3263, 3468 и 3497 кг / га, соответственно, по всем поперечным междурядьям и севооборотам. Усреднение по всем годам и севооборотам может быть статистически недостоверным, однако эти средние значения урожайности показывают, что 75- и 100% -ный уровень орошения имеет аналогичные значения урожайности и оба численно выше, чем 50% -ный уровень орошения.Таблица 3 также показывает, что значительная взаимосвязь урожайности действительно имела место между поливом севооборотом (2002) и поливом боковым поливом (2001). Не было взаимодействия урожайности между поливом, севооборотом и поперечным междурядьем.
В Таблице 3 значения вероятности показывают, что отводы капельниц, расположенные на расстоянии 1,83 м, имели такой же выход, как и отводы, расположенные на расстоянии 0,91 м, через девять из десяти лет. В 2000 г. была значительная разница в урожайности между двумя поперечными междурядьями.В течение вегетационного периода 2000 г. среднее расстояние между полями 0,91 м составляло 4028 кг / га −1 , в то время как поперечное расстояние 1,83 м составляло в среднем 2894 кг / га (Таблица 4). При усреднении по годам, поперечный интервал 0,91 м составлял в среднем 3484 кг / га, а поперечный интервал 1,83 м составлял в среднем 3334 кг / га. Преимущество более высокой урожайности в 150 кг / га при узком поперечном междурядье по сравнению с широким поперечным интервалом добавит к валовой выручке только около 68 долларов США / га. Широкое поперечное расстояние стоит около 377 долларов США / га только за трубку и вдвое больше — за узкое поперечное расстояние (754 доллара США / га по цене 0 долларов США.0672 / м НКТ). При таком уровне увеличения урожайности и выручки (150 кг / га и 68 долларов США / га соответственно) потребуется более 5,5 лет, чтобы оплатить стоимость только трубок, при условии, что арахис выращивался каждый год, а урожайность арахиса была статической. Могут быть понесены другие расходы, которые будут включать затраты на установку дополнительных фитинговых адаптеров, топливо, рабочую силу, техническое обслуживание и возможные другие критерии проектирования, такие как добавленные зоны, магистраль, клапаны и т. Д. Были значительные различия в доходности с поперечным взаимодействием вращения в трех из них. 9 лет (2000, 2002, 2006).Эти сельскохозяйственные годы также связаны с сильной реакцией на урожай либо в боковом (2000 г.), либо в севооборотном (2002 и 2006 гг.), Которые могут доминировать в этих реакциях взаимодействия. Следовательно, нет согласованности с взаимодействием «поперечное вращение», по которому можно было бы делать какие-либо долгосрочные выводы.
В целом, при использовании узкого поперечного междурядья повышение урожайности было незначительным, поэтому на этих почвах и в этом экологическом местоположении рекомендуется, чтобы боковые стволы располагались в чередующихся междурядьях для максимального урожая и возможной экономической отдачи.
Значения вероятности показывают, что севооборот значительно повлиял на урожайность арахиса в семь из восьми лет (Таблица 3). Во всех случаях, когда на урожайность существенно влиял севооборот, непрерывный арахис давал самый низкий урожай (Таблица 4). И наоборот, более высокие урожаи были измерены, когда периоды времени между посевами арахиса были длиннее. Хотя это и не является статистически достоверным, средняя урожайность при севообороте в течение периода времени проекта показала, что непрерывный арахис давал среднюю урожайность 2711 кг / га, в то время как севооборот хлопка-арахиса и кукурузы-арахиса через год составлял в среднем 3328 и 3651 кг / га соответственно.Двухлетние и трехлетние севообороты между культурами арахиса в среднем составляли 3912 и 4272 кг / га соответственно (см. Таблицу 4).
В течение срока реализации этого проекта севооборот кукурузы-арахиса через другой год имел тенденцию давать более высокий урожай арахиса по сравнению с севооборотом хлопка и арахиса через другой год. Севооборот кукуруза-арахис давал в среднем на 323 кг / га большую урожайность по сравнению с севооборотом хлопчатобумажно-арахисовый. Однако севооборот кукурузы и арахиса дает более высокие урожаи только в 50% случаев по сравнению с севооборотом хлопка и арахиса.Когда возникает ситуация по сокращению периода времени между севооборотом арахиса и чередованием года, производитель должен выбрать культуру (кукурузу или хлопок) с наилучшей экономической отдачей, а не с возможным увеличением урожайности арахиса.
Важность севооборота для арахиса известна уже много лет. Более низкий урожай арахиса при непрерывном производстве арахиса, вероятно, связан с повышенным давлением болезни (Henning et al ., 1982). Следовательно, рекомендуемый севооборот для получения максимального урожая арахиса будет заключаться в том, чтобы иметь как можно более длительные временные рамки между посевами арахиса, с особым упором на целостное планирование хозяйства для получения экономической отдачи. В целом, самый низкий TSMK наблюдался при непрерывном севообороте арахиса и увеличивался по мере увеличения времени между посевами арахиса.
Ирригационная обработка не оказала влияния на процентное содержание общего здорового зрелого ядра (TSMK) в течение года (Таблица 3).Боковое расстояние влияло на ЦМК два года из десяти, или 20% времени. Севооборот затрагивает ЦМК 7 из 8 лет или около 87% времени. Непрерывное вращение арахиса, как правило, имело самый низкий TSMK с более высоким процентом, возникающим по мере увеличения времени между вращениями арахиса. Наблюдалось значительное взаимодействие обработки для сортов и распределения зерен при поливе ротацией, поливе боковым, боковым вращением и поливом боковым вращением. Тем не менее, похоже, что нет какой-либо согласованности из года в год или согласованности отношений, чтобы делать какие-либо долгосрочные выводы на основе значимых взаимодействий.
В целом по мере увеличения TSMK процентный запас нефти имеет тенденцию к уменьшению. Орошение влияло на процентный запас масла в 20% случаев (Таблица 5). Поперечный интервал влияет на процентный запас масла только в 10% случаев, а севооборот влияет на запас масла в 62% случаев. За 10-летний период не было большой разницы в общем процентном соотношении ядер нефтяного сырья с обработкой орошением или поперечным интервалом.Однако севооборот оказал большое влияние на процентное содержание масличных культур. Непрерывные севообороты арахиса, как правило, имеют более высокий процент (более 7%) запаса масла по сравнению с другими ротациями. При севообороте арахиса в другой год процент запаса масла был таким же (6,4%) при усреднении за 10-летний период. Самый низкий процент запаса масла наблюдался при более длительных севооборотах, составляющих 3 (5,6%) и 4 (5,5%) лет между культурами арахиса (Таблица 5).
Таблица 5.Годовые значения содержания арахиса и среднее по проекту, состоящее из общего количества здоровых зрелых ядер (TSMK) и запаса масла для уровня орошения, поперечного расстояния и севооборота по годам с 1999 по 2007 год в Сассере, Джорджия.
Распределение ядер по размерам jumbos, medium и one не подвергалось ирригационной обработке. Боковое расстояние затрагивало ядра большого размера в 22% случаев, ядра среднего размера в 11% случаев и ядра первого размера в 22% случаев.Севооборот затрагивал jumbos, medium и one в 86%, 57% и 86% случаев соответственно. Обычно, когда процент jumbos уменьшается, процент медиумов и единиц увеличивается. По мере того, как уменьшались и джумбо, и медиумы, процент единиц увеличивался. Не было четких доказательств того, что какой-либо один севооборот влияет на распределение ядер по сравнению с другим, за исключением продолжающейся обработки арахиса. При такой непрерывной обработке арахиса крупные и средние ядра имели тенденцию к уменьшению, а ядра первого размера — к увеличению.Это означает, что более короткие периоды времени между посевами арахиса в севообороте могут отрицательно повлиять на размер ядра, но не обязательно на сорт арахиса или TSMK.
Использование глубокого подпочвенного капельного орошения возможно для арахиса и связанных с ним севооборотов. Средняя урожайность арахиса на протяжении всего срока реализации проекта показывает, что 75 и 100% уровень орошения имел сходные значения урожайности, и оба были численно выше, чем уровень орошения 50%.Кроме того, не наблюдалось снижения урожайности при внесении 75% рекомендованной воды по сравнению с полным (100%) рекомендованным количеством, что подразумевает возможную экономию воды на 25% при том же урожае.
Численное увеличение урожайности составило 150 кг / га с боковыми отводами, установленными под каждым рядком, по сравнению с альтернативными средними рядами. Однако валовая выручка, полученная от области, где трубки были размещены под каждым рядом, не может компенсировать затраты на дополнительные трубки по сравнению с промежуточными звеньями альтернативных рядов.Чтобы оплатить дополнительные трубы, потребуется более 5 лет постоянного увеличения добычи. Таким образом, рекомендуется, чтобы в этом ряду почв и условиях окружающей среды боковые стволы можно было размещать в чередующихся рядах для получения максимальной экономической урожайности и возможной экономической отдачи.
Не было четких доказательств в пользу кукурузы или хлопка до арахиса при чередовании года. Урожайность арахиса действительно увеличивалась по мере увеличения времени между посевами арахиса.Урожайность арахиса увеличилась на 27, 43 и 56% за 1, 2 и 3 года между посевами арахиса по сравнению с непрерывными посевами арахиса, соответственно. Для получения максимального урожая арахиса рекомендуется использовать как можно более длительные промежутки времени между посевами арахиса с упором на целостное планирование хозяйства для получения максимальной экономической отдачи.
На распределение зерна по размеру больше всего повлиял севооборот, а не норма полива или расстояние между ними.Обычно, когда процент jumbos уменьшался, процент медиумов и единиц увеличивался. Кроме того, когда уменьшались и jumbos, и медиумы, процент единиц увеличивался. Не было четких доказательств того, что какой-либо один севооборот отрицательно влияет на распределение ядер по сравнению с другим, за исключением продолжающейся обработки арахиса, когда ядра крупных и средних размеров имеют тенденцию к уменьшению, а ядра первого размера — к увеличению. Это означает, что более короткие периоды времени между посевами арахиса в севообороте могут отрицательно повлиять на размер ядра, но не обязательно на сорт арахиса или TSMK.
Боковое маскирование как функция интервала
Бэнкс, У. П. и Принцметал У. Конфигурационные эффекты при обработке визуальной информации. Восприятие и психофизика , 1976, 19 , 361–367.
Google Scholar
Бэнкс, У. П., Ларсон, Д. У., и Принцметал, У. Асимметрия визуальных помех. Восприятие и психофизика , 1979, 25 , 447–456.
Google Scholar
Бьорк, Э. Л., и Мюррей, Дж. Т. О природе входных каналов при визуальной обработке. Психологический обзор , 1977, 84 , 472–484.
Артикул PubMed Google Scholar
Баума, Х. Эффекты взаимодействия при распознавании парафовеальных букв. Nature , 1970, 226 , 177–178.
Артикул PubMed Google Scholar
Баума, Х.Визуальная интерференция при парафовеальном распознавании начальных и конечных букв слов. Исследование зрения , 1973, 13 , 767–782.
Артикул PubMed Google Scholar
Честейн Г. и Лоусон Л. Идентификация асимметрии парафовеальных пар стимулов. Восприятие и психофизика , 1979, 26 , 363–368.
Google Scholar
Эриксен, Б.A., & Eriksen, C. W. Влияние шумовых букв на идентификацию целевой буквы в непоисковой задаче. Восприятие и психофизика , 1974, 16 , 143–149.
Google Scholar
Эриксен, К. У. и Хоффман, Дж. Э. Степень обработки шумовых элементов во время выборочного кодирования с визуальных дисплеев. Восприятие и психофизика , 1973, 14 , 155–160.
Артикул Google Scholar
Эриксен, К.W., & Rohrbaugh, J. W. Некоторые факторы, определяющие эффективность избирательного внимания. Американский журнал психологии , 1970, 113 , 330–342.
Артикул Google Scholar
Эриксен, К. У., и Шульц, Д. У. {t Временные факторы в обработке визуальной информации}. В J. Requin (Ed.), Внимание и исполнение VII . Хиллсдейл, Нью-Джерси: Эрлбаум, 1978.
Google Scholar
Estes, W.К. Взаимодействие сигнальных и фоновых переменных при визуальной обработке. Восприятие и психофизика , 1972, 12 , 278–286.
Google Scholar
Флом, М. К., Уэймут, Ф. В., и Канеман, Д. Визуальное разрешение и взаимодействие контуров. Журнал Оптического общества Америки , 1963, 53 , 1026–1032.
Артикул PubMed Google Scholar
Фулд, К.Эффект сенсибилизации прямолинейными раздражителями. Vision Research , 1978, 18 , 1045–1051.
Артикул PubMed Google Scholar
Hubel, D. H., & Wiesel, T. N. Ferrier, лекция: Функциональная архитектура зрительной коры макак обезьяны. Труды Лондонского королевского общества (серия B), 1977, 198 , 1–59.
Артикул PubMed Google Scholar
Канеман, Д., & Хеник, А. Влияние визуальной группировки на немедленное вспоминание и избирательное внимание. В С. Дорник (ред.), Внимание и исполнение VI . Хиллсдейл, Нью-Джерси: Эрлбаум, 1977.
Google Scholar
Колерс П. А. и Рознер Б. С. О визуальной маскировке (метаконтраст): дихоптическое наблюдение. Американский журнал психологии , 1960, 73 , 2–21.
Артикул PubMed Google Scholar
Крумхансл, К.L. Наименование и расположение одновременно и последовательно представленных букв. Восприятие и психофизика , 1977, 22 , 293–302.
Артикул Google Scholar
Mackworth, N.H. Визуальный шум вызывает туннельное зрение. Психономическая наука , 1965, 3 , 67–68.
Google Scholar
Мэтьюз, М. Л. Локус презентации и эффект выборочной маскировки. Канадский журнал психологии , 1973, 27 , 343–349.
PubMed Google Scholar
Prinzmetal, W. Принципы интеграции функций в визуальном восприятии. Восприятие и психофизика , 1981, 30 , 330–340.
Артикул Google Scholar
Санти, Дж. Л. и Эгет, Х. Э. Независимость против вмешательства в перцептивную обработку букв. Восприятие и психофизика , 1982, 31 , 101–116.
Google Scholar
Treisman, A., & Schmidt, H. Иллюзорные союзы в восприятии объектов. Когнитивная психология , 1982, 14 , 107–141.
Артикул PubMed Google Scholar
Wolford, G. Модель возмущений для идентификации букв. Психологический обзор , 1975, 82 , 184–199.
Артикул PubMed Google Scholar
Вольффорд Г. и Холлингсворт С. Боковое маскирование при обработке визуальной информации. Восприятие и психофизика , 1974, 16 , 315–320.
Артикул Google Scholar
Цитокинин как позиционный сигнал, регулирующий расстояние между боковыми корнями у Arabidopsis | Журнал экспериментальной ботаники
Абстрактные
Корневые системы растений разработали адаптивную архитектуру для эксплуатации почвенных ресурсов.Формирование боковых корней (БК) способствует архитектуре корневой системы. Корни растений с более низким цитокининовым статусом образуют примордии LR (LRP) в необычно близкой близости, что указывает на роль гормона в регулировании расположения LR вдоль главной оси корня. Данные, полученные от мутантов по синтезу цитокинина Arabidopsis thaliana в сочетании с анализом экспрессии генов, показывают, что синтез цитокинина с помощью IPT5 и LOG4, происходящий на ранней стадии во время инициации LRP, генерирует локальный сигнал цитокинина, сокращающий образование LRP в соседних клетках перицикла.Кроме того, IPT3, IPT5 и IPT7 вносят вклад в синтез цитокининов в непосредственной близости от существующих LRP, тем самым подавляя инициацию новых LR. Интересно, что мутация генов CYP735A , необходимых для биосинтеза транс--зеатина, вызвала сильные дефекты в позиционировании LR, указывая на важную роль этого метаболита цитокинина в регулировании расстояния LR. Кроме того, показано, что цитокинин и известный регулятор расстояния LR, рецептор-подобная киназа ARABIDOPSIS CRINKLY4 (ACR4), действуют неиерархическим образом, но могут осуществлять реципрокный контроль на уровне транскриптов.Взятые вместе, результаты показывают, что цитокинин действует как паракринный гормональный сигнал, регулирующий архитектуру корневой системы.
Введение
Боковые корни (БК) составляют основную часть корневой системы растений и важны для оптимизации способности корневой системы усваивать питательные вещества и воду из почвы. В Arabidopsis LR образуются вдоль главной оси корня из клеток перицикла (ПК) полюса ксилемы в соответствии с определенными закономерностями, такими как то, что зачатки боковых корней (LRP) обычно не образуются рядом или напротив друг друга (De Smet et al. al., 2006; Дубровский и др. , 2006; рассмотрено Dastidar et al. , 2012). Xylem pole PCs онтогенетически примированы, чтобы формировать компетентные сайты для образования LR вдоль продольной оси. Этот предварительный паттерн устанавливается с помощью эндогенного часового механизма, названного LR-часами (Moreno-Risueno et al. , 2010). Однако не все компетентные клетки развиваются в LRP или LRs и механизмы, формирующие паттерн локального пространственного расположения LRP, остаются в значительной степени не охарактеризованными. Похоже, что ауксин играет роль в регулировании расстояния между двумя последовательными LRP через модуль передачи сигналов BDL / IAA12-MP⁄ARF5 (De Smet et al., 2010) и регуляторы транскрипции, названные PLETHORA3 (PLT3), PLT5 и PLT7 (Hofhuis et al. , 2013). Более того, рецептор-подобная киназа Arabidopsis CRINKLY4 (ACR4), как сообщается, действует вне клеток автономно, чтобы предотвратить инициацию бокового корня (LRI) соседних ПК (De Smet et al. , 2008). Была выдвинута гипотеза, что члены семейства сигнальных пептидов GOLVEN (GLV) / ROOT Growth FACTORS / CLE-LIKE взаимодействуют с ACR4, чтобы инициировать сигнальный каскад (Meng et al., 2012; Fernandez et al. , 2013).
Цитокинины представляют собой другой класс растительных гормонов, играющих решающую роль в регуляции образования и роста LR, действующих как негативный регулятор ранних процессов, предотвращая установление градиента ауксина, необходимого для образования LR (Laplaze et al. , 2007; Marhavý и др. , 2011). Метаболизм цитокининов и сигнальные пути A. thaliana были выяснены и включают ~ 60 генов (обзор Sakakibara, 2006; Werner and Schmülling, 2009; Kieber and Schaller, 2014).Вкратце, первая и лимитирующая стадия биосинтеза цитокинина катализируется изопентенилтрансферазами (IPT), а последующее образование метаболитов транс--зеатина ( t Z) из метаболитов изопентениладенина (iP) катализируется гидроксилазой. ферменты (цитохром P450 CYP735A), кодируемые двумя преимущественно корневыми генами (Takei et al. , 2004 a ; Kiba et al. , 2013). Заключительным этапом синтеза цитокининов iP- и t Z-типа является высвобождение активных цитокининов из их нуклеотидных предшественников с помощью цитокининовых нуклеозид-5′-монофосфатфосфорибогидролаз, называемых LONELY GUY (LOG) (Kurakawa et al., 2007). Распад цитокининов катализируется за одну стадию ферментами цитокининоксидаза / дегидрогеназа (CKX) (Werner et al. , 2006). Все эти метаболические ферменты кодируются в Arabidopsis семействами генов из нескольких членов. Сигнал цитокинина воспринимается через три мембранных рецептора гистидинкиназы (названных AHK2, AHK3 и CRE1 / AHK4) (Inoue et al. , 2001; Yamada et al. , 2001) и передается фосфотрансмиттерными белками (AHPs). ) (Hutchison et al., 2006) в ядро, где регуляторы ответа типа B (ARR типа B) активируют нижестоящие гены, определяющие действие цитокининов (Mason et al. , 2005). В Arabidopsis один член семейства AHP, AHP6, лишен канонического фосфо-акцептирующего остатка His и действует как негативный регулятор передачи сигналов цитокининов (Mähönen et al. , 2006).
Первые намеки на то, что цитокинин может обеспечивать позиционную информацию для LR, были получены из анализа цитокинин-дефицитных трансгенных растений CKX , которые показали дефекты в расстоянии LR (Werner et al., 2003). Кроме того, более высокая плотность LR неоднократно отмечалась на растениях с более низким содержанием цитокининов или передачей сигналов (Werner et al. , 2003, 2010; Mason et al. , 2005; Miyawaki et al. , 2006; Riefler и др. , 2006; Laplaze и др. , 2007; Heyl и др. , 2008; Bielach и др. , 2012; Chang и др. , 2013). Более конкретно, локальная эктопическая экспрессия гена CKX в клетках-основателях боковых корней (LRFC) вызвала более высокую плотность LR.Это указывает на то, что LRFCs могут быть источником цитокининового сигнала (Laplaze et al. , 2007), который может восприниматься в зонах высокой чувствительности к цитокининам вблизи LRP (Bielach et al. , 2012). Здесь роль цитокинина в контроле позиционирования LR была проанализирована более подробно и идентифицированы соответствующие гены метаболизма цитокининов и передачи сигналов. Предполагается, что ингибирующие концентрации цитокинина происходят внутри и вокруг существующего LRP, и для их создания необходимы гены IPT и LOG4 .Также обсуждается взаимосвязь между этим путем и путем ACR4 в контроле ризотаксиса.
Материалы и методы
Растительный материал и условия роста
Все растения A. thaliana , использованные в этом исследовании, относились к экотипу Колумбия (Col-0). Семена acr4 ( acr4-2 , acr4-2 crr2-2 acr4-2 crr4 ; De Smet et al. , 2008), ipt ( ipt3-2 , ipt5-2 , ipt7-1 , ipt3-2 ipt5-3 , ipt3-2 ipt7-1, ipt3-2 ipt5-3 ipt7-1 ; Miyawaki et al., 2006), log4-3 (Kuroha et al. , 2009) и cyp735a1-1, cyp735a2-2, cyp735a1-1 cyp735a2-2 (Kiba et al. , 2013) мутанты были любезно предоставлены Ивом Де Сметом, Тацуо Какимото и Хитоши Сакакибара соответственно. Маркерные генные линии pCKX: GUS (Werner et al. , 2003), pIPT: GFP (Takei et al. , 2004 b ), pLOG: GUS (Kuroha et al. , 2009), pCYP735A2: GUS (Kiba et al., 2013), ACR4 :: h3B: YFP (Гиффорд и др. , 2003), CycB1; 1: GUS (Бекман и др. , 2001), TCSn: GFP (Zürcher et al., , 2013) и DR5: GUS (Ulmasov et al. , 1997) были получены от Hitoshi Sakakibara, Gwyneth C. Ingram, Laurent Laplaze, Bruno Müller и Nottingham Stock Centre Arabidopsis соответственно. 35S: CKX1 , мутантные линии гена типа B ARR ( arr1-3 arr10-5 , arr1-3 arr11-2 , arr1-3 arr12-1 , arr10-5 arr12- 1 , arr11-2 arr12-1 ), и двойные мутанты рецептора цитокинина ahk2-5 ahk3-7 , ahk2-5 cre1-2 и ahk3-7 cre1-2 были описаны ранее ( Werner et al., 2003; Mason et al. , 2005; Riefler et al. , 2006).
Семена стерилизовали поверхность и высевали на твердую среду (среда Мурашиге и Скуга [MS] половинной концентрации, 1% сахароза и 0,9% агар, pH 5,7) и стратифицировали при 4 ° C в течение 3 дней в темноте перед прорастанием. После стратификации все планшеты помещали вертикально в белый свет (~ 100 мкмоль м -2 с -1 ) в условиях длинного дня (16 часов света / 8 часов темноты) при 22 ° C. Для обработки цитокинином проростки проращивали и выращивали на среде MS половинной концентрации с 6-бензиламинопурином (BA) или без него в течение 11 дней.
Микроскопические анализы
Гистохимическое окрашивание на активность β-глюкуронидазы (GUS) проводили в основном в соответствии с Jefferson et al. (1987) с изменениями Hemerly et al. (1993). Длина первичного корня измерялась на цифровых изображениях пластин с использованием программного обеспечения ImageJ 1.40 (http://rsb.info.nih.gov/ij/). Для регистрации картины окрашивания GUS и организации LRP образцы очищали и устанавливали в соответствии с Malamy and Benfey (1997).Все образцы анализировали методом дифференциальной интерференционной контрастной микроскопии (Zeiss Axioskop). Количество ПК между соседними LRP подсчитывали в направлении отростка, начиная с LRP, ближайшего к кончику первичного корня.
Для флуоресцентной микроскопии проростки помещали в воду под покровным стеклом для анализа сигналов желтого флуоресцентного белка (YFP) или зеленого флуоресцентного белка (GFP) с использованием конфокального лазерного сканирующего микроскопа Leica TCS SP5. Для слитого белка YFP длина волны 517 нм и фильтр 526–568 нм использовались для возбуждения и анализа сигналов соответственно.Сигнал флуоресценции GFP анализировали с помощью аргонового лазера 488 нм в сочетании с набором фильтров 500–530 нм.
Количественная ПЦР с обратной транскрипцией в реальном времени
Суммарную РНК экстрагировали из корней растений Arabidopsis возрастом 11 дней с использованием метода TRIzol, как описано Brenner et al. (2005). Синтез кДНК, количественная ПЦР (qPCR) и анализ данных были выполнены согласно Werner et al. (2010). UBC10 (At5g53300) и PDF1 (At3g25800) использовали в качестве эталонных генов для нормализации уровней экспрессии.Последовательности праймеров для всех проанализированных генов перечислены в дополнительной таблице S1.
Результаты
Позиционирование LRP изменено в цитокинин-дефицитных растениях
Во-первых, расстояние LR в цитокинин-дефицитных проростках 35S: CKX1 трансгенных Arabidopsis (Werner et al. , 2003) сравнивали с мутантными проростками acr4 , которые, как известно, имеют дефект позиционирования LR (и др. , 2008) и проростки дикого типа в одинаковых условиях роста.В корнях дикого типа почти нет LRP или появившихся LR, инициированных непосредственно из соседних или противоположных сайтов (Рис. 1A, B). Среди 373 событий LRI в 15 корнях дикого типа только пять LRP (1,3 ± 0,6%) были расположены в непосредственной близости (рис. 1J). Напротив, у acr4 мутантных растений LRP инициируется из соседних или противоположных сайтов (Fig. 1C, D), что согласуется с предыдущими результатами (De Smet et al. , 2008). Всего 39 LRP из 348 событий LRI (11,0 ± 1,9%) были сформированы из этих аберрантных положений (рис.1J). Точно так же 76 LRP из 811 событий LRI (9,1 ± 0,9%) были расположены непосредственно рядом или напротив друг друга у растений 35S: CKX1 (рис. 1J). Наблюдалась фенотипическая изменчивость дефектного паттерна расстановки. В некоторых случаях участки нескольких PCs показали активность клеточного деления (Fig. 1E) и близко расположенные LRP формировались либо на продольной оси (Fig. 1F), либо на противоположной стороне оси (Fig. 1G). Близко расположенные LRP в основном принадлежали к одной и той же стадии развития или различались только одной стадией (согласно Malamy and Benfey, 1997), что указывает на то, что они индуцировались в одно и то же время (рис.1F, G). Хотя и реже, растения с дефицитом цитокинина также иногда образовывали кластеры близко расположенных LRP на разных стадиях роста (Fig. 1H). Редко, несколько близко расположенных LRP появлялись и появлялись как кластерные LRs (Fig. 1I). Однако доля возникших соседних LR была низкой, это указывает на то, что большая часть LRP в аберрантных позициях останавливается во время развития и не вносит большого вклада в общую корневую архитектуру. Примечательно, что первичные корни мутантов acr4 были только примерно на 15% короче, чем корни дикого типа (данные не показаны), а корни трансгенных растений 35S: CKX1 были еще длиннее (Werner et al., 2003), указывая на то, что более короткое расстояние между LRP не коррелирует с длиной первичного корня.
Рис. 1.
Расстояние LR изменено у растений с дефицитом цитокинина. (A-I) Никаких проксимальных LRP и LR не наблюдали у дикого типа (A, B), тогда как LRP и LR в непосредственной близости наблюдались у acr4 (C, D) и 35S: CKX1 (E-I). Красные звездочки указывают границы LRP или появившихся LR. (J) Доля LRP, разделенных различным количеством PC в 11-дневных растениях дикого типа, acr4 и дефицитных по цитокинину 35S: CKX1 растений.n (количество проанализированных корней) = 15. (K) Экспрессия CycB1; 1: GUS и маркерные гены DR5: GUS указывают на деление соседних PC в растениях 35S: CKX1 , но не в растениях дикого типа и нормальный образец ауксина в аберрантно расположенных LRP и LR. Черные стрелки указывают на LRP. Размер стержня в (A-I) и (K) составляет 50 мкМ. Достоверность различий в (J) анализировали с помощью двустороннего критерия Стьюдента t . * P <0,05; ** P <0.01; *** P <0,001. Планки погрешностей указывают на SEM (эта цифра доступна в цвете по адресу JXB онлайн).
Рис. 1.
Расстояние LR изменено у растений с дефицитом цитокинина. (A-I) Никаких проксимальных LRP и LR не наблюдали у дикого типа (A, B), тогда как LRP и LR в непосредственной близости наблюдались у acr4 (C, D) и 35S: CKX1 (E-I). Красные звездочки указывают границы LRP или появившихся LR. (J) Доля LRP, разделенных различным количеством PC в 11-дневных растениях дикого типа, acr4 и дефицитных по цитокинину 35S: CKX1 растений.n (количество проанализированных корней) = 15. (K) Экспрессия CycB1; 1: GUS и маркерные гены DR5: GUS указывают на деление соседних PC в растениях 35S: CKX1 , но не в растениях дикого типа и нормальный образец ауксина в аберрантно расположенных LRP и LR. Черные стрелки указывают на LRP. Размер стержня в (A-I) и (K) составляет 50 мкМ. Достоверность различий в (J) анализировали с помощью двустороннего критерия Стьюдента t . * P <0,05; ** P <0.01; *** P <0,001. Планки погрешностей указывают на SEM (эта цифра доступна в цвете по адресу JXB онлайн).
Чтобы изучить возможность дефектного позиционирования LRP, процент LRP, разделенных нулем, одним или максимум тремя ПК, был определен в корнях растений дикого типа, acr4 и 35S: CKX1 (рис. 1J). ). Наибольшая разница между диким типом и мутантами была обнаружена для расстояния в ноль ПК, то есть непосредственно соседних клеток, и повышенная частота также наблюдалась у мутантов для расстояния LRP до трех ПК (рис.1J). У дикого типа всего 21,5% LRP имели соседний LRP на этом расстоянии, в то время как это было 48,0% и 50,0% для acr4 и 35S: CKX1 , соответственно. Все остальные LRP имели соседний LRP на большем расстоянии. Вместе дефекты расстояния LR у мутантов acr4 и 35S: CKX1 растений встречались с одинаковой частотой и с аналогичным пространственным распределением.
Для анализа активности деления клеток во время образования LRP, экспрессия репортерного маркера CycB1; 1: GUS , делящего LRFC (Casimoro et al., 2001) сравнивали между растениями дикого типа и растениями с дефицитом цитокинина. У дикого типа окрашивание GUS наблюдалось только в LRFC, но не в соседних PC. Напротив, в цитокинин-дефицитных корнях соседние PCs могут быть активированы одновременно, чтобы стать LRFCs, на что указывает активность клеточного деления после их спецификации (Fig. 1K). Чтобы исследовать, устанавливают ли близко расположенные LRP нормальный ауксиновый паттерн, репортер ауксина DR5: GUS анализировали на фоне 35S: CKX1 . DR5: GUS маркирует клетки-основатели и последующее образование зачатков как в LRP дикого типа, так и в соседних LRP у 35S: CKX1 растений и демонстрирует сходные паттерны экспрессии (фиг. 1K). По-видимому, близкорасположенные LRP устанавливают нормальное клеточное деление и паттерн ауксина и лишены дефектов роста, наблюдаемых у др. Мутантов, демонстрирующих пространственно измененный LRI (Hofhuis et al. , 2013).
Гены метаболизма и передачи сигналов цитокининов участвуют в позиционировании LRP
Фенотип расстояния LR у цитокинин-дефицитных растений предполагает, что обычно происходящее ингибирование образования LRP в непосредственной близости от LRP, которое начало формироваться, отменено.Для изучения пространственной организации генерации сигнала, опосредующей это ингибирование, были проанализированы паттерны экспрессии и функции генов синтеза цитокининов. Сообщалось, что экспрессия нескольких генов синтеза цитокининов, как IPT , так и LOG , а также расщепляющих цитокинин генов CKX , происходит во время образования LR (Werner et al. , 2003; Miyawaki et al. др. , 2004; Takei и др. , 2004b; Kuroha и др., 2009 г .; Parizot et al. , 2010; Kiba et al. , 2013). Пространственно-временной паттерн экспрессии этих генов на разных стадиях образования LR был изучен более подробно, потому что в большинстве случаев сообщалось только об одной или двух стадиях. Особенно любопытный паттерн экспрессии был продемонстрирован генами IPT5 , LOG4 и CKX6 , которые были включены очень рано во время образования LRP на стадии I, аналогично DR5: GUS (рисунки 1K и 2; дополнительные Инжир.S1). Во время дальнейшего развития LRP IPT5 и LOG4 экспрессировались в большинстве клеток до появления. В появившихся LR экспрессия IPT5 стала ограничиваться кончиком корня, а LOG4 сильно экспрессировался в проваскулярной ткани и меристеме LR (фиг. 2). Напротив, экспрессия CKX6 наблюдалась в сосудистой сети и только на стадии I LRP, но не на следующих стадиях (дополнительные данные). Репортер pIPT3: GFP и pIPT7: GFP не проявлял активности во время раннего развития LR. IPT3 экспрессировался в PCs и базальной стеле молодых LRs, IPT7: экспрессия GFP ограничивалась преимущественно сосудистой стелой первичного корня (Fig. 2). Экспрессия репортерных генов для LOG3 , CKX1 и CKX5 стала видимой только в сосудистой сети и в основе возникшего LR (дополнительный рис. S1). Также для pCYP735A2: GUS не наблюдалось экспрессии в самом зачатке, но наблюдались различные паттерны и сила экспрессии в ПК и сосудистой ткани по обе стороны от существующих зачатков (рис.2). Паттерн экспрессии pCYP735A1: GUS не мог быть проанализирован из-за его низкого уровня экспрессии (Takei et al. , 2004 a ; Kiba et al. , 2013). Анализ экспрессии генов показал специфический паттерн экспрессии генов синтеза цитокининов и указал на гены IPT5 и LOG4 как на, возможно, особенно важные для создания источника цитокинина в молодых зачатках. Их экспрессия может устанавливать ингибирующее поле, предотвращающее LRI в соседних клетках.
Рис. 2.
Пространственно-временная экспрессия выбранных генов синтеза цитокининов во время развития LR. Последовательность развития паттерна экспрессии промоторов генов метаболизма цитокининов показана слева направо, начиная с стадии I LRP до появившейся LR. Соответствующий промоутер указан в верхнем левом углу каждой серии изображений. Трехдневные проростки окрашивали реакционным буфером GUS в течение 1 часа и очищали. Экспрессию GFP анализировали в 5-дневных проростках с помощью конфокального лазерного сканирующего микроскопа.Часть представленного здесь паттерна окрашивания корней, сообщающего об экспрессии генов метаболизма цитокининов, была опубликована ранее (Werner et al. , 2003; Takei et al. , 2004 b ; Kuroha et al. , 2009; Киба и др. , 2013). Масштабная линейка составляет 50 мкМ (эта цифра доступна в цвете по ссылке JXB онлайн).
Рис. 2.
Пространственно-временная экспрессия выбранных генов синтеза цитокининов во время развития LR. Последовательность развития паттерна экспрессии промоторов генов метаболизма цитокининов показана слева направо, начиная с стадии I LRP до появившейся LR.Соответствующий промоутер указан в верхнем левом углу каждой серии изображений. Трехдневные проростки окрашивали реакционным буфером GUS в течение 1 часа и очищали. Экспрессию GFP анализировали в 5-дневных проростках с помощью конфокального лазерного сканирующего микроскопа. Часть представленного здесь паттерна окрашивания корней, сообщающего об экспрессии генов метаболизма цитокининов, была опубликована ранее (Werner et al. , 2003; Takei et al. , 2004 b ; Kuroha et al. , 2009; Kiba et al., 2013). Масштабная линейка составляет 50 мкМ (эта цифра доступна в цвете по ссылке JXB онлайн).
Для дальнейшего изучения роли генов синтеза цитокининов в позиционировании LR был проведен подробный анализ с использованием мутантов одного и более высокого порядка. Одиночные мутанты ipt показали только слабые ( ipt3 , ipt5 ) или отсутствующие ( ipt7 ) дефекты позиционирования LRP (фиг. 3A). Двойные мутанты ipt3 5 и ipt3 7 показали повышенную долю аномально расположенных LRP по сравнению с диким типом, но только разница, наблюдаемая для ipt3 5 , была статистически значимой.Доля смещенных LRP увеличилась до 13,6 ± 2,0% (57 из 416 LRP в 12 корнях) в ipt3 5 7 корнях (фиг. 3A). Неуместные LR в мутантах ipt демонстрируют паттерн, аналогичный показанному в растениях 35S: CKX1 , и включают различные стадии развития от LRP до появляющихся LR (дополнительный рис. S2). Мутант log4 также обнаруживал дефекты расстояния LR (7,0 ± 1,4%) (фиг. 3A).
Рис. 3.
Снижение содержания цитокининов или передачи сигналов увеличивает частоту дефектов позиционирования LRP.(A) Процент LRP, расположенных непосредственно рядом друг с другом в 11-дневных проростках мутантов по синтезу цитокинина. n (количество проанализированных корней) = 12–16. (B) Доля LRP, разделенных от нуля до трех ПК в диком типе (WT; n = 30) и мутантах по синтезу цитокинина ipt3 5 7 и log4 . n = 15–17. (C) Процент LRP, расположенных непосредственно рядом друг с другом у 11-дневных проростков мутантов, передающих сигнал цитокинину. n = 11–14. Достоверность различий в (A-C) анализировали с помощью двустороннего критерия Стьюдента t .* P <0,05; ** P <0,01; *** P <0,001. Планки погрешностей указывают на SEM.
Рис. 3.
Снижение содержания цитокининов или передачи сигналов увеличивает частоту дефектов позиционирования LRP. (A) Процент LRP, расположенных непосредственно рядом друг с другом в 11-дневных проростках мутантов по синтезу цитокинина. n (количество проанализированных корней) = 12–16. (B) Доля LRP, разделенных от нуля до трех ПК в диком типе (WT; n = 30) и мутантах по синтезу цитокинина ipt3 5 7 и log4 .n = 15–17. (C) Процент LRP, расположенных непосредственно рядом друг с другом у 11-дневных проростков мутантов, передающих сигнал цитокинину. n = 11–14. Достоверность различий в (A-C) анализировали с помощью двустороннего критерия Стьюдента t . * P <0,05; ** P <0,01; *** P <0,001. Планки погрешностей указывают на SEM.
Одинарные и двойные мутанты генов CYP735A также показали фенотип аномального позиционирования LRP. Одинарные мутантные фенотипы cyp735a1 и cyp735a2 были усилены у двойного мутанта cyp735a1 a2 с 13.9 ± 3,0% событий LRI расположены в непосредственной близости (рис. 3A). Этот двойной мутант содержал менее 5% уровней – цитокининов Z-типа дикого типа, но в 2 раза более высокий уровень цитокининов iP-типа (Kiba et al. , 2013). Таким образом, мутантный фенотип указывает на особую функциональную значимость t Z в регуляции спейсинга LRP.
Фенотип дефекта спейсинга LRP был дополнительно проанализирован путем подсчета количества ПК между двумя близко расположенными LRP в корнях ipt3 5 7 и log4 по сравнению с диким типом.На фиг. 3В показано, что оба мутанта по синтезу цитокинина имели более высокую долю LRP, разделенных от нуля до трех ПК (44,5% для ipt3 5 7 и 47,9% для log4 ) по сравнению с диким типом (22,8%). Наибольшая разница в спецификации LRP между мутантами по синтезу цитокинина и диким типом была непосредственно рядом с уже инициированным LR, а затем постепенно уменьшалась, показывая почти одинаковую частоту на расстоянии трех PC (рис. 3B). Этот градиент активности, возможно, может быть связан с потоком цитокининов, возникающим в молодых LRP и вызванным комбинированной активностью IPT5 и LOG4, которые в первую очередь влияют на непосредственно соседние клетки (рис.2А). Рис. 4 показывает, что сниженный синтез цитокининов внутри и вокруг LRP влияет на выходной сигнал цитокинина в зоне высокой чувствительности к цитокининам, окружающей LRP (Bielach et al. , 2012). Сигнал репортера цитокинина TCSn: GFP (Zürcher et al. , 2013) у дикого типа был высоким в клетках, соседних с LRP, но ниже в этих клетках в корнях мутанта по синтезу цитокинина ipt3 5 7 . Это согласуется с более низкой ингибирующей активностью цитокинина в отношении инициации LR в этих клетках.
Рис. 4.
Экспрессия TCSn: GFP во время разработки LR. Активность цитокинина, видимая с помощью TCSn: GFP , высока в PC по обе стороны от LRP у растений дикого типа. TCSn: экспрессия GFP снижена в ПК, прилегающих к LRP цитокинин-дефицитного мутанта ipt3 5 7 . Белые звездочки обозначают границы LRP или появившихся LR. Масштабная шкала составляет 100 мкМ (эта цифра доступна в цвете на JXB в Интернете).
Рис. 4.
Экспрессия TCSn: GFP во время разработки LR. Активность цитокинина, видимая с помощью TCSn: GFP , высока в PC по обе стороны от LRP у растений дикого типа. TCSn: экспрессия GFP снижена в ПК, прилегающих к LRP цитокинин-дефицитного мутанта ipt3 5 7 . Белые звездочки обозначают границы LRP или появившихся LR. Масштабная шкала составляет 100 мкМ (эта цифра доступна в цвете на JXB в Интернете).
Чтобы исследовать, участвуют ли гены рецептора цитокинина и гены регулятора ответа в регуляции расстояния LR, были проанализированы мутанты двойного рецептора и мутанты ARR типа B .Более высокий процент неуместных LRP по сравнению с диким типом был сформирован во всех трех двойных мутантах рецептора, но разница не была статистически значимой (фиг. 3C). Среди генов-регуляторов ответа типа B, двойные мутанты arr1 12, (6,1 ± 1,6%) и arr10 12, (6,8 ± 2,2%) показали значительно увеличенные дефекты спейсинга по сравнению с диким типом (рис. 3C). Эти данные согласуются с высокой степенью функциональной избыточности рецепторов цитокининов и генов типа B ARR в регуляции развития LR (Chang et al., 2013) и демонстрируют, что восприятие и передача сигнала цитокинина важны для регулирования расстояния LR.
Экспрессия ACR4 снижена в корнях цитокинин-дефицитных растений, а статус цитокинина понижен у мутантов acr4Поскольку и цитокининовые, и ACR4 пути предотвращают LRI соседних PC, была исследована взаимосвязь между этими путями. Проанализирована экспрессия ACR4 в корнях 11-дневных проростков мутантов, синтезирующих и сигнализирующих цитокинин.На фиг. 5А показано, что количество транскриптов ACR4 было ниже в корнях большинства растений с пониженным цитокининовым статусом. Наиболее сильное снижение количества транскриптов ACR4 (~ 60% снижение по сравнению с диким типом) было отмечено у тройных мутантов ipt3 5 7 . Кроме того, у дикого типа, вызванная pACR4 экспрессия h3B: YFP наблюдалась в LRP, тогда как флуоресценция YFP с трудом определялась на фоне дефицита цитокинина 35S: CKX1 и ipt3 5 7 (фиг.5Б). Однако анализ транскрипта ACR4 в разные моменты времени после экзогенного добавления цитокинина не выявил сильного влияния цитокинина на устойчивый уровень мРНК ACR4 (Brenner et al. , 2012 и данные не показаны). Взятые вместе, это показывает, что экспрессия ACR4 снижена в цитокинин-дефицитных растениях, но что ACR4 не является первичным геном цитокининового ответа. Уменьшение расстояния LR в цитокинин-дефицитных растениях может частично достигаться за счет снижения экспрессии ACR4 .
Рис. 5.
Взаимодействие между сигнальными путями цитокинина и ACR4 на уровне экспрессии генов. (A) Анализ экспрессии ACR4 с помощью кПЦР в корнях 11-дневных различных проростков с дефицитом цитокинина. (B) Экспрессия ACR4 :: h3B: YFP (зеленые сигналы) видна в LRP дикого типа (WT), но отсутствует у растений с дефицитом цитокинина. Масштабная шкала составляет 20 мкМ. (C) Анализ экспрессии гена GLV с помощью кПЦР в корнях 11-дневных различных проростков с дефицитом цитокинина.(D) Профили транскриптов метаболизма цитокининов и сигнальных генов в корнях 11-дневных одинарных и двойных мутантов acr4 . Планки погрешностей представляют собой SEM из трех (A) или двух (C, D) биологических повторов. Каждая биологическая копия содержала корни по крайней мере шести отдельных растений. Во всех случаях уровень экспрессии дикого типа был установлен на 1, и статистическая значимость различий значений экспрессии у мутантов по сравнению с диким типом определялась с помощью теста Стьюдента t (* P <0.05) (этот рисунок доступен в цвете по ссылке JXB онлайн).
Рис. 5.
Взаимодействие между сигнальными путями цитокинина и ACR4 на уровне экспрессии генов. (A) Анализ экспрессии ACR4 с помощью кПЦР в корнях 11-дневных различных проростков с дефицитом цитокинина. (B) Экспрессия ACR4 :: h3B: YFP (зеленые сигналы) видна в LRP дикого типа (WT), но отсутствует у растений с дефицитом цитокинина. Масштабная шкала составляет 20 мкМ. (C) Анализ экспрессии гена GLV с помощью кПЦР в корнях 11-дневных различных проростков с дефицитом цитокинина.(D) Профили транскриптов метаболизма цитокининов и сигнальных генов в корнях 11-дневных одинарных и двойных мутантов acr4 . Планки погрешностей представляют собой SEM из трех (A) или двух (C, D) биологических повторов. Каждая биологическая копия содержала корни по крайней мере шести отдельных растений. Во всех случаях уровень экспрессии дикого типа был установлен на 1, и статистическая значимость различий значений экспрессии у мутантов по сравнению с диким типом определялась с помощью теста Стьюдента t (* P <0.05) (этот рисунок доступен в цвете по ссылке JXB онлайн).
Затем были исследованы уровни транскриптов генов GLV , которые кодируют небольшие сигнальные пептиды, которые являются предполагаемыми лигандами ACR4 (Meng et al. , 2012; Fernandez et al. , 2013). Экспрессия всех пяти генов GLV , которые специфически экспрессируются на ранних стадиях развития LR (Fernandez et al. , 2013), показала более низкую экспрессию в корнях растений с глобально сниженным цитокининовым статусом (рис.5C), хотя они образуют больше LRP. Самое сильное снижение уровней транскрипта гена GLV (~ 70–80% по сравнению с диким типом) снова наблюдалось у трех мутантных растений ipt3 5 7 , аналогично сильному снижению ACR4 (рис. 5A, C). ). Однако, напротив, двойной мутант cyp735a и мутант log4 не демонстрировали сильных изменений экспрессии гена GLV по сравнению с диким типом. Возможно, изменения в t Z не влияют на экспрессию гена GLV и что локальные изменения содержания цитокининов, ожидаемые в мутантах log4 , разводятся в экстрактах цельного корня.В любом случае цитокинин, по-видимому, положительно регулирует экспрессию гена GLV в корнях. Однако еще предстоит выяснить, приводит ли это к изменению активности ACR4, которая влияет на LRI.
Профили транскриптов генов цитокининов были проанализированы с помощью кПЦР в корнях мутантов acr4 и в двойных мутантах, сочетающих acr4 с мутантными аллелями двух других членов семейства, CRR2 и CRR4 (De Smet et al. др. , 2008 г.). Анализ показал значительно изменившиеся уровни транскриптов для нескольких генов цитокининов. IPT3 и IPT5 оба были подавлены в мутантах acr4 , тогда как CKX1 и CKX6 были активированы (фиг. 5D). Точно так же несколько генов, кодирующих факторы передачи сигналов цитокинина, были подавлены, включая ген рецептора AHK3 и гены типа B ARR ARR1 и ARR12 . В совокупности измененная экспрессия генов цитокининов предполагает более низкий статус цитокининов в мутантных корнях acr4 .Интересно, что CYP735A2 был сильно активирован в двойных мутантах acr4 ccr2 и acr4 crr4 , что указывает на отрицательный контроль системы ACR4 на синтез цитокининов t Z-типа (рис. 5D).
Цитокинин и ACR4 ингибируют инициирование соседних ПК через независимые неиерархические пути
Для дальнейшего исследования того, контролируют ли цитокинин и ACR4 позиционирование LR одним и тем же или разными путями, фенотип LR-интервала гибридных проростков acr4 35S: CKX1 сравнивали с родительскими фенотипами.Гибридные растения acr4 35S: CKX1 показали гораздо более высокий процент аберрантно расположенных LRP (20,0 ± 2,9%), чем у обоих родителей (фиг. 6A). Этот в значительной степени аддитивный эффект предполагает, что цитокинин и ACR4 действуют, по крайней мере частично, отдельными путями, предотвращая соседние PC от LRI. Соответственно, добавление цитокинина к питательной среде оказало лишь слабое влияние на дефект расстояния LR у мутантов acr4 , тогда как оно уменьшило дефект расстояния LR на 67% у проростков 35S: CKX1 (рис.6Б).
Рис. 6.
Эффекты пониженного цитокининового статуса и мутации ACR4 на расстояние LR являются аддитивными. (A) Процент непосредственно прилегающих LRP в 11-дневных корнях дикого типа (WT), 35S: CKX1 , acr4 и acr4 35S: CKX1 гибридов. n = 15–18. (B) Неправильная замена LRP была частично дополнена экзогенным цитокинином (BA) в 35S: CKX1 растениях, но не в acr4 . Перед анализом проростки выращивали в течение 11 дней на среде с 20 нМ БА или без нее.n = 15 для дикого типа, 32 для acr4 и 15 для 35S: CKX1 . Планки погрешностей в (A, B) указывают на SEM. Статистическая значимость различий была рассчитана с использованием двустороннего критерия Стьюдента t (** P <0,01; *** P <0,001).
Рис. 6.
Эффекты пониженного цитокининового статуса и мутации ACR4 на расстояние LR являются аддитивными. (A) Процент непосредственно прилегающих LRP в 11-дневных корнях дикого типа (WT), 35S: CKX1 , acr4 и acr4 35S: CKX1 гибридов.n = 15–18. (B) Неправильная замена LRP была частично дополнена экзогенным цитокинином (BA) в 35S: CKX1 растениях, но не в acr4 . Перед анализом проростки выращивали в течение 11 дней на среде с 20 нМ БА или без нее. n = 15 для дикого типа, 32 для acr4 и 15 для 35S: CKX1 . Планки погрешностей в (A, B) указывают на SEM. Статистическая значимость различий была рассчитана с использованием двустороннего критерия Стьюдента t (** P <0,01; *** P <0.001).
Обсуждение
Расположение LRP вдоль главного корня Arabidopsis определяет обычно регулярный ризотаксический паттерн, и LRP не образуются в непосредственной близости друг от друга. Гормон цитокинин действует в подавлении образования LRP, близкого к уже существующим, и, таким образом, действует как позиционный сигнал, регулирующий расстояние между LRP. Позиционный сигнал генерируется путем точной настройки экспрессии генов синтеза цитокининов, контролируемой локально и с точки зрения развития.Предполагается, что меньшее расстояние LRP у мутантов, синтезирующих цитокинин, связано с локальным ингибирующим градиентом цитокинина, происходящим от LRFC, и с цитокинином, продуцируемым в PC, соседних с LRFC. Это паракринное действие цитокинина, комбинация не клеточно-автономной и клеточно-автономной активности, ингибирует образование LRP в ПК, соседствующих с существующими LRP. Эта гипотеза подтверждается (i) активацией генов синтеза цитокининов на ранней стадии развития LRP и их экспрессией в соседних PC; (ii) тот факт, что нарушение этих генов вызывает аберрантное позиционирование LRP, с самым сильным эффектом в непосредственной близости от существующего LRP; и (iii) постепенное снижение ингибирующего действия цитокининов у дикого типа с увеличением расстояния от LRFC.Предполагаемая активность цитокинина суммирована в модели (рис. 7).
Рис. 7.
Модель, иллюстрирующая действие цитокинина в регулировании расстояния LR. Модель предсказывает, что подавление LRI в ПК, соседствующих с существующими LRFC, является комбинированным эффектом цитокинина, синтезируемого в LRFC, и цитокинина, синтезируемого соседними клетками. Активность IPT5 и LOG4 в LRFCs вызывает образование iP, который распространяется латерально. Соседние клетки конвертируют iP в t Z с помощью CYP735A2 и, кроме того, синтезируют свой собственный цитокинин для ингибирования образования LR.LRFC характеризуются высоким отношением ауксина к цитокинину, обозначенным оранжевым цветом, в то время как соседние ПК характеризуются низким отношением ауксина к цитокинину, обозначенным зеленым (эта цифра доступна в цвете на JXB онлайн) .
Рис. 7.
Модель, иллюстрирующая действие цитокинина в регулировании расстояния LR. Модель предсказывает, что подавление LRI в ПК, соседствующих с существующими LRFC, является комбинированным эффектом цитокинина, синтезируемого в LRFC, и цитокинина, синтезируемого соседними клетками.Активность IPT5 и LOG4 в LRFCs вызывает образование iP, который распространяется латерально. Соседние клетки конвертируют iP в t Z с помощью CYP735A2 и, кроме того, синтезируют свой собственный цитокинин для ингибирования образования LR. LRFC характеризуются высоким отношением ауксина к цитокинину, обозначенным оранжевым цветом, в то время как соседние ПК характеризуются низким отношением ауксина к цитокинину, обозначенным зеленым (эта цифра доступна в цвете на JXB онлайн) .
Характер экспрессии генов синтеза цитокининов показал, что IPT5 и LOG4 экспрессировались очень рано во время инициации LR (рис.2) (Miyawaki et al. , 2004; Kuroha et al. , 2009). Однако экспрессия репортера TCS: GFP показала, что передача сигналов цитокинина сильно репрессируется на ранних фазах развития LR (Bielach et al. , 2012), что указывает на то, что сигнал, генерируемый IPT5 и LOG4 в инициированном LR, подавлялся или рассеивался. сбоку. Учитывая сильное усиление аномального позиционирования LR при мутации IPT5 в дополнение к IPT3 и IPT7 и, в частности, дефект позиционирования LR, вызванный одной только мутацией LOG4 (рис.3A, B), возможно, что цитокинин, синтезируемый IPT5 и LOG4 в LRFC, передается на соседние ПК для предотвращения LRI. Это согласуется с самой низкой частотой новых LRP, непосредственно соседствующих с существующими, у дикого типа и их постепенно увеличивающейся частотой с увеличением расстояния от существующих LRP (Fig. 3B). Соответственно, наиболее сильное увеличение аберрантно расположенных LRP у мутантов ipt3 5 7 и log4 было обнаружено непосредственно рядом с существующими мутантами (рис.3Б). Кроме того, экспрессия расщепляющего цитокинин гена CKX в LRFC снижает расстояние от нового соседнего LRP, что соответствует подавляющей функции цитокинина, производного от LRFC, в соседних клетках (Laplaze et al. , 2007). В соответствии с идеей цитокининового сигнала, исходящего от LRFCs, было обнаружено, что выходной сигнал цитокинина обнаруживается через несколько часов после LRI в соседних клетках (Bielach et al. , 2012). Однако авторы не сообщали о перемещении цитокининового сигнала от клетки к клетке.Альтернативной возможностью объяснить постепенное различие в ответе ПК, соседних с LRFC, на подавляющий сигнал цитокинина, может быть различие в их чувствительности к цитокинину. Однако сильно увеличенный процент LRP, непосредственно прилегающего к существующему LRP, не обнаружен у мутантов, сохраняющих одиночные цитокининовые рецепторы (Fig. 3C), хотя они обычно обнаруживают более высокую плотность LRP и LR (Chang et al. , 2013). В любом случае существует дополнительная значимая роль генов синтеза цитокининов ( IPT3 , IPT7 ), экспрессируемых в ткани перицикла, прилегающей к существующей LRP.Это свидетельствует о комбинированном действии цитокинина, производного от LRP, и цитокинина, синтезируемого в соседних клетках (рис. 7).
Недавно сообщалось о других случаях действия цитокининов на короткое расстояние с участием LOG4. Было показано, что продукция цитокинина LOG4 в эмбриональных клетках-предшественниках ксилемы индуцирует пролиферацию прокамбиальных клеток в соседних клетках, тем самым регулируя рост и формирование паттерна во время развития сосудов не-клеточно-автономным образом (De Rybel et al. , 2014; Ohashi- Ито и др., 2014). Кроме того, вычислительная модель продемонстрировала, что цитокинин, генерируемый LOG4, может передавать сигналы по нескольким клеточным слоям в апикальной меристеме побегов и действовать вместе с передачей сигналов CLV как позиционный сигнал в нише стволовых клеток (Chikarmane et al. , 2012). Это указывает на то, что последняя стадия активации во время биосинтеза цитокининов играет ключевую роль в установлении локальных градиентов цитокининов и, более конкретно, предполагает, что LOG4 играет важную роль в формировании клеточного паттерна.
Анализ мутантов cyp735a показал, что t Z обладает сильной активностью в подавлении образования LRP. Это раскрывает новую локальную функцию генов CYP735A , известную своей важностью в синтезе t цитокинина Z-типа для коммуникации от корня к побегу (Kiba et al. , 2013). CYP735A2 Активность репортерного гена, управляемая промотором , присутствовала в клетках, окружающих LRP, но не в самой LRP. CYP735A2 — один из генов, наиболее сильно подавляемых ауксином ARF7 / ARF19-зависимым образом в корне (Parizot et al., 2010 г.). К сожалению, из-за низкой экспрессии CYP735A1 не было аналогичной информации для оценки его поведения в этом контексте. Однако может случиться так, что iP, синтезируемый IPT5 и LOG4 в LRFC, достигает соседних клеток путем диффузии и / или транспорта, а затем превращается в t Z.
Каким образом активность цитокинина и регуляция гена синтеза цитокинина могут быть связаны к ауксину, который является главным регулятором развития LR? Некоторые признаки получены из исследований экспрессии генов.Экспрессия IPT5 активируется во время образования LR ARF7 / ARF19-зависимым образом (Parizot et al. , 2010), и было показано, что промотор LOG4 является прямой мишенью для факторов транскрипции TMO5-LHW. действует ниже модуля ответа на ауксин BDL / IAA12-MP / ARF5 (De Rybel et al. , 2014), действуя немного позже, чем ARF7-ARF19 в LRI (De Smet et al. , 2010). Интересно, что мутант mp обнаруживает дефект расстояния LR (De Smet et al., 2010), что может быть частично связано с уменьшением экспрессии LOG4 . Дополнительные гены цитокининов, которые также включаются посредством путей ARF7-ARF19 и TMO5-LHW, представляют собой два ауксин-зависимых гена CKX ( CKX1 , CKX6 ) (Parizot et al. , 2010) и auxin- ответный негативный регулятор AHP6 (Ohashi-Ito et al. , 2014), который экспрессируется в LR с ранних стадий и далее (Moreira et al. , 2013).Активация этих генов в LRFCs может поддерживать низкий цитокининовый статус в этих клетках, чтобы избежать негативного влияния цитокинина на их дальнейшее развитие. В самом деле, случайное деление периклинальных клеток в LRFC мутантов ahp6 указывает на репрессивное действие AHP6 на передачу сигналов цитокинина в этих клетках (Moreira et al. , 2013). Взятые вместе, ауксин-индуцированное снижение статуса цитокинина в LRFC и синтез цитокинина в соседних клетках будет способствовать высокому соотношению ауксин / низкий цитокинин в LRFC и низкому соотношению ауксин / высокий цитокинин в соседних клетках.Сравнение репортерных сигналов DR5 и TCS неизменно показывает, что максимумы ответа ауксина и цитокинина комплементарны в перицикле и во время развития LR (Benková et al. , 2003; Bielach et al. , 2012).
Сильно пониженный статус цитокининов не снимает ингибирования всех PCs полюса ксилемы, которые онтогенетически примированы, чтобы в конечном итоге стать LRFCs, предполагая, что одна или несколько других систем работают параллельно, чтобы контролировать расстояние LR.Одна из таких систем основана на ACR4. Экспрессия ACR4 была ниже в корнях цитокинин-дефицитных растений, а мутанты acr4 имели более низкий цитокининовый статус (фиг. 5). Генетический анализ не выявил иерархического порядка между цитокинином и путем ACR4 (рис. 6). Однако реципрокное влияние на уровни транскриптов указывает на существование перекрестных помех и предполагает, что ACR4 оказывает положительное влияние на путь цитокининов и наоборот . Это взаимное влияние могло быть постепенным и не обязательно сильно влиять на результат генетического анализа, хотя процент неуместных LRP был выше, чем это было бы в результате аддитивного эффекта у гибридов acr4 35S: CKX1 (рис.6А). Более того, степень местного реципрокного влияния на уровни транскриптов не могла быть оценена, поскольку они были измерены в экстрактах цельного корня. В любом случае, один путь может действовать посредством регуляции транскрипции с прямой связью, чтобы усилить подавление LRI другим путем в соседних клетках.
В совокупности результаты показывают участие цитокинина в регуляции архитектуры корневой системы вскоре после отбора клеток-основателей, первого ключевого шага в образовании LR из примированной клетки.Предполагается, что цитокинин действует локально как паракринный сигнал, который исходит из LRFC и соседних с ним клеток, действуя одновременно как супрессор LRI и определяя, какие из компетентных сайтов развиваются в LR. Поучительный сигнал от LRFC к соседним компьютерам для LRI сэкономит ресурсы и поможет разработать оптимальную корневую систему. Кроме того, цитокинин может действовать как эндогенный сигнал, реагируя на сигналы окружающей среды, такие как питательные вещества и осмотические условия (Malamy et al. , 2005; Ramireddy et al., 2014). Таким образом, он способствует развитию пластичности корневой архитектуры, которая должна постоянно адаптироваться к очень сложной подземной среде.
Сокращения:
LR
LRP
LRFC
Ячейка-основатель бокового корня
PC
BA6
7
- t
Мы благодарим докторов Hitoshi Sakakibara, Ive De Smet, Tatsuo Kakimoto, Gwyneth C.Инграм, Лоран Лаплаз и Бруно Мюллер; Ноттингемский фондовый центр арабидопсиса (NASC) за предоставление семенного материала; и д-ра Томаша Вернера за критическое прочтение рукописи. Эта работа была поддержана финансированием Федерального министерства науки и технологий (BMBF) в рамках проекта ROOT (грант 0315704 A) и Далемского центра растениеводства (DCPS), приоритетной области FU Berlin.
Список литературы
Бикман
т
Бурссенс
S
Inzé
D
.2001
.Периклеточный цикл Arabidopsis
.Журнал экспериментальной ботаники
52
,403
—411
.Benková
E
Michniewicz
м
Зауэр
м
Тейхманн
т
Seifertová
D
Юргенс
G
Фримл
Дж
.2003
.Локальные, зависимые от оттока градиенты ауксина как общий модуль для формирования растительных органов
.Ячейка
115
,591
—602
.Белях
А
Podlešáková
К
Marhavý
-П
Duclercq
Дж
Куэста
С
Мюллер
В
Грюневальд
Вт
Тарковски
-П
Benková
E
.2012
.Пространственно-временная регуляция органогенеза боковых корней у арабидопсиса цитокинином
.Заводская ячейка
24
,3967
—3981
.Бреннер
WG
Романов
GA
Кёльмер
Я
Bürkle
л
Schmülling
т
.2005
.Гены немедленного и замедленного цитокининового ответа Arabidopsis thaliana , идентифицированные с помощью полногеномного профилирования экспрессии, выявляют новые чувствительные к цитокинину процессы и предполагают действие цитокининов через транскрипционные каскады
.Заводской журнал
44
,314
—333
.Бреннер
WG
Рамиредди
E
Хейл
А
Schmülling
т
.2012
.Регуляция гена цитокинином в Arabidopsis
.Границы растениеводства
3
,8
.Казимиро
Я
Марчант
А
Бхалерао
RP
и другие. .2001
.Транспорт ауксина способствует инициации бокового корня Arabidopsis
.Заводская ячейка
13
,843
—852
.Чанг
л
Рамиредди
E
Schmülling
т
.2013
.Образование и рост боковых корней Arabidopsis избыточно регулируется метаболизмом цитокининов и сигнальными генами
.Журнал экспериментальной ботаники
64
,5021
—5032
.Курица
VS
Гордон
SP
Tarr
PT
Heisler
MG
Мейеровиц
EM
.2012
.Передача сигналов цитокинина как позиционный сигнал для формирования паттерна апикально-базальной оси растущей меристемы побега Arabidopsis
.Proceedings of the National Academy of Sciences, USA
109
,4002
—4007
.Дастидар
MG
Жуанне
В
Майзель
А
.2012
.Корневое ветвление: механизмы, прочность, пластичность
.Междисциплинарные обзоры Wiley: биология развития
1
,329
—343
.Де Рыбель
В
Адиби
м
Бреда
AS
и другие. .2014
.Интеграция роста и формирования паттерна во время формирования сосудистой ткани у Arabidopsis
.Наука
345
,1255215
.Де Смет
Я
Ваннест
S
Inzé
D
Бикман
т
.2006
.Зарождение бокового корня или рождение новой меристемы
.Молекулярная биология растений
60
,871
—887
.Де Смет
Я
Василева
В
Де Рыбель
В
и другие. .2008
.Рецептороподобная киназа ACR4 ограничивает деления формирующихся клеток в корне
Arabidopsis .Наука
322
,594
—597
.Де Смет
Я
Lau
S
Voß
U
и другие. .2010
.Бимодульный ответ на ауксин контролирует органогенез у Arabidopsis
.Proceedings of the National Academy of Sciences, USA
107
,2705
—2710
.Дубровский
JG
Гамбетта
GA
Эрнандес-Баррера
А
Шишкова
S
Гонсалес
Я
.2006
.Зарождение бокового корня у Arabidopsis : окно развития, пространственный паттерн, плотность и предсказуемость
.Анналы ботаники
97
,903
—915
.Фернандес
А
Drozdzecki
А
Hoogewijs
К
Нгуен
А
Бикман
т
Марена
А
Хилсон
-П
.2013
.Транскрипционная и функциональная классификация сигнальных пептидов GOLVEN / ROOT Growth FACTOR / CLE-подобных показывает их роль в формировании боковых корней и волос
.Физиология растений
161
,954
—970
.Гиффорд
ML
Декан
S
Инграм
GC
.2003
.Ген Arabidopsis ACR4 играет роль в организации клеточного слоя во время развития покровов семяпочки и краев чашелистика
.Девелопмент
130
,4249
—4258
.Хемерли
AS
Феррейра
-П
de Almeida Engler
Дж
Ван Монтегю
м
Энглер
G
Inzé
D
.1993
. Экспрессияcdc2a в Arabidopsis связана с компетенцией к делению клеток
.Заводская ячейка
5
,1711
—1723
.Хейл
А
Рамиредди
E
Бреннер
WG
Рифлер
м
Allemeersch
Дж
Schmülling
т
.2008
.Репрессор транскрипции ARR1-SRDX подавляет активность плейотропных цитокининов у Arabidopsis
.Физиология растений
147
,1380
—1395
.Hofhuis
H
Ласковски
м
Ду
Я
Прасад
К
Григг
S
Пинон
В
Шерес
В
.2013
.Филлотаксис и ризотаксис в Arabidopsis модифицированы тремя транскрипционными факторами PLETHORA
.Текущая биология
23
,956
—962
.Hutchison
CE
Ли
Дж
Аргуэсо
С
и другие. .2006
.Белки фосфотрансфера гистидина Arabidopsis являются избыточными позитивными регуляторами передачи сигналов цитокининов
.Plant Cell
18
,3073
—3087
.Иноуэ
т
Хигучи
м
Хашимото
Я
Секи
м
Кобаяши
м
Като
т
Табата
S
Шинозаки
К
Какимото
т
.2001
.Идентификация CRE1 как рецептора цитокинина из Arabidopsis
.Природа
409
,1060
—1063
.Джефферсон
RA
Кавана
TA
Беван
МВт
.1987
.GUS слияния: бета-глюкуронидаза как чувствительный и универсальный маркер слияния генов у высших растений
.Журнал EMBO
6
,3901
—3907
.Киба
т
Такеи
К
Кодзима
м
Сакакибара
H
.2013
.Модификация боковой цепи цитокининов контролирует рост побегов Arabidopsis
.Клетка развития
27
,452
—461
.Кибер
JJ
Шаллер
GE
.2014
.Цитокинины
.The Arabidopsis Book
,e0168
.Куракава
т
Уэда
N
Маэкава
м
Кобаяши
К
Кодзима
м
Нагато
Я
Сакакибара
H
Кёдзука
Дж
.2007
.Прямой контроль активности меристемы побегов цитокинин-активирующим ферментом
.Природа
445
,652
—655
.Куроха
т
Tokunaga
H
Кодзима
м
Уэда
N
Исида
т
Нагава
S
Фукуда
H
Сугимото
К
Сакакибара
H
.2009
.Функциональный анализ цитокинин-активирующих ферментов LONELY GUY показывает важность прямого пути активации у Arabidopsis
.Заводская ячейка
21
,3152
—3169
.Laplaze
л
Бенкова
E
Казимиро
Я
и другие. .2007
.Цитокинины действуют непосредственно на клетки-основатели бокового корня, подавляя инициацию корня
.Заводская ячейка
19
,3889
—3900
.Mähönen
AP
Хигучи
м
Тормакангас
К
Мияваки
К
Pischke
MS
Sussman
MR
Helariutta
Я
Какимото
т
.2006
.Цитокинины регулируют двунаправленную фосфорелейную сеть в Arabidopsis
.Текущая биология
16
,1116
—1122
.Маламы
JE
Бенфей
PN
.1997
.Организация и дифференцировка клеток в боковых корнях Arabidopsis thaliana
.Девелопмент
124
,33
—44
.Маламы
JE
.2005
.Внутренние пути реакции и реакции окружающей среды, которые регулируют архитектуру корневой системы
.Завод, клетки и окружающая среда
28
,67
—77
.Marhavý
-П
Белях
А
Абас
л
и другие. .2011
.Цитокинин модулирует эндоцитарный перенос переносчика оттока ауксина PIN1 для контроля органогенеза растений
.Клетка развития
21
,796
—804
.Мейсон
MG
Мэтьюз
DE
Аргирос
DA
Максвелл
BB
Кибер
JJ
Алонсо
JM
Эккер
JR
Шаллер
GE
.2005
.Множественные регуляторы ответа типа B опосредуют передачу сигнала цитокинина в Arabidopsis
.Заводская ячейка
17
,3007
—3018
.Мэн
л
Бьюкенен
BB
Фельдман
LJ
Луан
S
.2012
.CLE-подобные (CLEL) пептиды контролируют характер роста корней и развитие боковых корней у Arabidopsis
.Proceedings of the National Academy of Sciences, USA
109
,1760
—1765
.Мияваки
К
Мацумото-Китано
м
Какимото
т
.2004
.Экспрессия генов изопентенилтрансфераз, биосинтезирующих цитокинины, в Arabidopsis : тканевая специфичность и регуляция ауксином, цитокинином и нитратом
.Заводской журнал
37
,128
—138
.Мияваки
К
Тарковски
-П
Мацумото-Китано
м
Като
т
Сато
S
Тарковская
D
Табата
S
Сандберг
G
Какимото
т
.2006
.Роль Arabidopsis АТФ / АДФ изопентенилтрансфераз и тРНК изопентенилтрансфераз в биосинтезе цитокининов
.Proceedings of the National Academy of Sciences, USA
103
,16598
—16603
.Морейра
S
Бишопп
А
Карвалью
H
Campilho
А
.2013
.AHP6 Ингибирует передачу сигналов цитокинина для регулирования ориентации деления клеток перицикла во время инициации бокового корня
.PloS One
8
,e56370
.Морено-Рисуено
MA
Ван Норман
JM
Морено
А
Чжан
Дж
Анерт
SE
Бенфей
PN
.2010
.Осциллирующая экспрессия гена определяет способность к периодическому ветвлению корня Arabidopsis
.Наука
329
,1306
—1311
.Охаши-Ито
К
Саегуса
м
Ивамото
К
Oda
Я
Катаяма
H
Кодзима
м
Сакакибара
H
Фукуда
H
.2014
.Комплекс bHLH активирует деление сосудистых клеток за счет действия цитокининов в апикальной меристеме корня
.Текущая биология
24
,2053
—2058
.Паризот
В
Де Рыбель
В
Бикман
т
.2010
.VisuaLRTC: новый взгляд на инициацию бокового корня путем объединения определенных наборов данных транскриптомов
.Физиология растений
153
,34
—40
.Рамиредди
E
Чанг
л
Schmülling
т
.2014
.Цитокинин как посредник для регулирования архитектуры корневой системы в ответ на сигналы окружающей среды
.Сигнализация и поведение предприятия
9
,e27771
.Рифлер
м
Новак
O
Strnad
м
Schmülling
т
.2006
.Мутанты рецептора цитокининов Arabidopsis обнаруживают функции роста побегов, старения листьев, размера семян, прорастания, развития корней и метаболизма цитокининов
.Заводская ячейка
18
,40
—54
.Сакакибара
H
.2006
.Цитокинины: активность, биосинтез и транслокация
.Ежегодный обзор биологии растений
57
,431
—449
.Такеи
К
Ямая
т
Сакакибара
H
.2004
а .Arabidopsis CYP735A1 и CYP735A2 кодируют гидроксилазы цитокинина, которые катализируют биосинтез транс-зеатина
.Журнал биологической химии
279
,41866
—41872
.Такеи
К
Уэда
N
Аоки
К
Куромори
т
Хираяма
т
Шинозаки
К
Ямая
т
Сакакибара
H
.2004
б .AtIPT3 является ключевым детерминантом нитрат-зависимого биосинтеза цитокининов у Arabidopsis
.Физиология растений и клеток
45
,1053
—1062
.Ульмасов
т
Мерфетт
Дж
Hagen
G
Гильфойл
ТДж
.1997
.Белки Aux / IAA подавляют экспрессию репортерных генов, содержащих природные и высокоактивные синтетические элементы ответа на ауксин
.Заводская ячейка
9
,1963
—1971
.Вернер
т
Кёльмер
Я
Бартрина
Я
Холст
К
Schmülling
т
.2006
.Новое понимание биологии деградации цитокининов
.Биология растений
8
,371
—381
.Вернер
т
Мотыка
В
Лауку
В
Смец
R
Ван Онкелен
H
Schmülling
т
.2003
.Цитокинин-дефицитные трансгенные растения Arabidopsis демонстрируют множественные изменения в развитии, указывающие на противоположные функции цитокининов в регуляции активности меристем побегов и корней
.Заводская ячейка
15
,2532
—2550
.Вернер
т
Nehnevajova
E
Кёльмер
Я
Новак
O
Strnad
м
Крамер
U
Schmülling
т
.2010
.Специфичное для корней снижение цитокинина вызывает усиление роста корней, засухоустойчивость и обогащение листьев минералами у Arabidopsis и Tobacco
.Plant Cell
22
,3905
—3920
.Вернер
т
Schmülling
т
.2009
.Действие цитокининов в развитии растений
.Текущее мнение по биологии растений
12
,527
—538
.Ямада
H
Сузуки
т
Терада
К
Такеи
К
Исикава
К
Miwa
К
Ямашино
т
Mizuno
т
.2001
.Гистидинкиназа Arabidopsis AHK4 представляет собой цитокинин-связывающий рецептор, который передает сигналы цитокинина через мембрану
.Физиология растений и клеток
42
,1017
—1023
.Zürcher
E
Тавор-Деслекс
D
Литуев
D
Энкерли
К
Tarr
PT
Мюллер
В
.2013
.Надежный и чувствительный синтетический датчик для мониторинга транскрипционного выхода сигнальной сети цитокининов в planta
.Физиология растений
161
,1066
—1075
.© Автор, 2015. Опубликовано издательством Oxford University Press от имени Общества экспериментальной биологии.
Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License (http: // creativecommons.org / licenses / by / 3.0 /), который разрешает неограниченное повторное использование, распространение и воспроизведение на любом носителе при условии правильного цитирования оригинальной работы.Капельное орошение Поперечный интервал и мульчирование влияют на схему увлажнения, регулирование роста корней и урожай кукурузы в песчаной почве
Автор
Включено в список:- Чжоу, Лифенг
- Фэн, Хао
- Чжао, Ин
- Ци, Чжицзюань
- Чжан, Тибин
- Хэ, Цзяньцян
- Дайк, Майлз
Abstract
Почва с слоем песка толщиной 20–60 см (60–90 см ниже поверхности) была недавно рекультивирована для использования ее потенциала производства зерна в засушливом регионе на северо-западе Китая.Двухлетнее полевое исследование было проведено в ирригационном районе Хетао для изучения влияния бокового зазора и методов мульчирования почвы при капельном орошении на влажность почвы, NO3−, регулирование роста побегов и эффективность использования воды (WUE) яровой кукурузы. . Коэффициент однородности Кристиансена (Cus) был принят для оценки влажности почвы и распределения NO3–, которое рассчитывалось с учетом содержания влаги в почве и концентрации NO3–. Четыре изученных варианта обработки состояли из двух поливов с боковыми интервалами (A1: 1.0 м, A2: 0,5 м) и два режима покрытия пленкой (M1: полностью мульчированный, M2: частично мульчированный).
Предлагаемое цитирование
- Чжоу, Лифэн и Фэн, Хао и Чжао, Ин и Ци, Чжицзюань и Чжан, Тибин и Хэ, Цзяньцян и Дайк, Майлз, 2017. « Поперечный интервал капельного орошения и мульчирование влияют на схему увлажнения, регулирование роста побегов и урожай кукурузы в песчаной почве », Управление водными ресурсами в сельском хозяйстве, Elsevier, vol. 184 (C), страницы 114-123.
Обозначение: RePEc: eee: agiwat: v: 184: y: 2017: i: c: p: 114-123
DOI: 10.1016 / j.agwat.2017.01.008Скачать полный текст от издателя
Поскольку доступ к этому документу ограничен, вы можете поискать его другую версию.
Ссылки на IDEAS
- Эль-Хендави, Салах Э. и Шмидхальтер, Урс, 2010 г. « Оптимальные сочетания между частотой полива и нормой для капельного орошения кукурузы, выращиваемой на песчаной почве ,» Управление водными ресурсами в сельском хозяйстве, Elsevier, vol.97 (3), страницы 439-448, март.
- Эль-Хендави, Салах Э. и Эль-Латтиф, Эссам А. Абд и Ахмед, Мохамед С. и Шмидхальтер, Урс, 2008. « Влияние скорости орошения и плотности растений на урожайность и эффективность водопользования кукурузы для капельного орошения ,» Управление водными ресурсами в сельском хозяйстве, Elsevier, vol. 95 (7), страницы 836-844, июль.
- Паломо, М. Дж., Морено, Ф., Фернандес, Дж. Э., Диас-Эспехо, А., Хирон, И. Ф., 2002. « Определение потребления воды в оливковых садах с использованием подхода водного баланса ,» Управление водными ресурсами в сельском хозяйстве, Elsevier, vol.55 (1), страницы 15-35, май.
- Ли, Цзюшен и Чжан, Цзяньцзюнь и Рао, Миньцзе, 2004 г. « Характер увлажнения и распределение азота в зависимости от стратегии фертигации из точечного источника на поверхности », Управление водными ресурсами в сельском хозяйстве, Elsevier, vol. 67 (2), страницы 89-104, июнь.
- Фэн, Чжао-Чжун и Ван, Сяо-Кэ и Фэн, Цзун-Вэй, 2005. « Выщелачивание азота и засоления почвы после осеннего орошения и его влияние на грунтовые воды в ирригационном районе Хетао, Китай », Управление водными ресурсами в сельском хозяйстве, Elsevier, vol.71 (2), страницы 131-143, февраль.
- Michelakis, N. & Vougioucalou, E. & Clapaki, G., 1993. « Использование воды, объем увлажненной почвы, распределение корней и урожай авокадо при капельном орошении ,» Управление водными ресурсами в сельском хозяйстве, Elsevier, vol. 24 (2), страницы 119–131, октябрь.
- Аль-Огайди, Ахмед А.М. И Ваяйок, Эмрун и Роушон, М.К. И Абдулла, Ахмед Фикри, 2016. « Оценка характера увлажнения в системах капельного орошения с использованием улучшенной эмпирической модели », Управление водными ресурсами в сельском хозяйстве, Elsevier, vol.176 (C), страницы 203-213.
- Zotarelli, L. & Dukes, M.D. & Scholberg, J.M.S. И Муньос-Карпена, Р. и Исерман, Дж., 2009. « Накопление азота в томатах и эффективность использования удобрений на песчаной почве в зависимости от нормы азота и графика полива ,» Управление водными ресурсами в сельском хозяйстве, Elsevier, vol. 96 (8), страницы 1247-1258, август.
- Bozkurt, Yesim & Yazar, Attila & Gencel, Burcin & Sezen, Metin Semih, 2006. « Оптимальное расстояние по бокам для кукурузы с капельным орошением в Средиземноморском регионе Турции ,» Управление водными ресурсами в сельском хозяйстве, Elsevier, vol.85 (1-2), страницы 113-120, сентябрь.
- Ван, Цзыкуй и Ву, Путе и Чжао, Синин и Гао, Ин и Чен, Сяоли, 2015. « Коэффициент водопользования и урожайности в системе посева полос пшеницы и кукурузы для засушливого региона на северо-западе Китая ,» Управление водными ресурсами в сельском хозяйстве, Elsevier, vol. 161 (C), страницы 77-85.
Цитаты
Цитаты извлекаются проектом CitEc, подпишитесь на его RSS-канал для этого элемента.
Цитируется:- Wang, Dan & Mo, Yan & Li, Guangyong & Wilkerson, Carol Jo & Hoogenboom, Gerrit, 2021. « Повышение урожайности кукурузы и уменьшение остатков азота в почве с использованием мульчированного капельного фертигации », Управление водными ресурсами в сельском хозяйстве, Elsevier, vol. 251 (С).
- Zou, Haiyang & Fan, Junliang & Zhang, Fucang & Xiang, Youzhen & Wu, Lifeng & Yan, Shicheng, 2020. « Оптимизация режимов капельного орошения и внесения удобрений для повышения урожайности зерна, продуктивности воды сельскохозяйственных культур и экономических выгод от яровой кукурузы в Северо-Западном Китае », Управление водными ресурсами в сельском хозяйстве, Elsevier, vol.230 (С).
- Шмидт, Дженнифер Э. и Петерсон, Кейтлин и Ван, Даоюань и Скоу, Кейт М. и Годен, Амели К.М., 2018. « Компромиссы Агроэкосистемы, связанные с переходом на подземное капельное орошение в органических системах », Управление водными ресурсами в сельском хозяйстве, Elsevier, vol. 202 (C), страницы 1-8.
- Чжоу, Лифэн и Хэ, Цзяньцян и Ци, Чжицзюань и Дайк, Майлз и Цзоу, Юфэн и Чжан, Тибин и Фэн, Хао, 2018. « Влияние бокового расстояния для капельного орошения и мульчирования на распределение воды и нитратов в почве, урожай кукурузы и эффективность водопользования ,» Управление водными ресурсами в сельском хозяйстве, Elsevier, vol.199 (C), страницы 190-200.
- Wang, Feng & Xiao, Junfu & Ming, Bo & Xie, Ruizhi & Wang, Keru & Hou, Peng & Liu, Guangzhou & Zhang, Guoqiang & Chen, Jianglu & Liu, Wanmao & Yang, Yunshan & Qin, Anzhen & Ли, Шаокун, 2021 год. « Урожайность зерна и динамика эвапотранспирации кукурузы на капельном орошении при высокой плотности посадки в засушливом и полувлажном климате », Управление водными ресурсами в сельском хозяйстве, Elsevier, vol. 247 (С).
Самые популярные товары
Это элементы, которые чаще всего цитируют те же работы, что и эта, и цитируются в тех же работах, что и эта.- Коуту, А., Руис Падин, А., Рейносо, Б., 2013. « Сравнительная урожайность и эффективность использования воды двух гибридов кукурузы, различающихся по зрелости при спринклерном спринклерном режиме и двух системах капельного орошения с разными боковыми интервалами в Леоне, Испания ,» Управление водными ресурсами в сельском хозяйстве, Elsevier, vol. 124 (C), страницы 77-84.
- Чжоу, Лифэн и Хэ, Цзяньцян и Ци, Чжицзюань и Дайк, Майлз и Цзоу, Юфэн и Чжан, Тибин и Фэн, Хао, 2018. « Влияние бокового расстояния для капельного орошения и мульчирования на распределение воды и нитратов в почве, урожай кукурузы и эффективность водопользования ,» Управление водными ресурсами в сельском хозяйстве, Elsevier, vol.199 (C), страницы 190-200.
- Lv, Чжаоянь и Диао, Мин и Ли, Вэйхуа и Цай, Цзянь и Чжоу, Цинь и Ван, Сяо и Дай, Тинбо и Цао, Вэйсин и Цзян, Донг, 2019. « Влияние поперечного расстояния на пространственные различия в использовании воды и урожайности яровой пшеницы в разных рядах системы капельного орошения », Управление водными ресурсами в сельском хозяйстве, Elsevier, vol. 212 (C), страницы 252-261.
- Мерли, Кэмерон Б. и Шарма, Сумит и Уоррен, Джейсон Г.И Арналл, Дэрил Б. и Раун, Уильям Р., 2018. « Реакция урожайности кукурузы и зерна сорго на смещения рядов на подпочвенных отводах ,» Управление водными ресурсами в сельском хозяйстве, Elsevier, vol. 208 (C), страницы 357-362.
- Аттиа, Ахмед и Эль-Хендави, Салах и Аль-Сухайбани, Насер и Алотаиби, Маджед и Тахир, Мухаммад Усман и Камаль, Халед Ю., 2021. « Оценка стратегии планирования недостаточного орошения для повышения урожайности и продуктивности воды кукурузы в засушливых условиях с использованием моделирования », Управление водными ресурсами в сельском хозяйстве, Elsevier, vol.249 (С).
- Мухаммад Зайн и Чжуанюн Си, Сен Ли и Ян Гао, Фейсал Мехмуд и Шафик-Ур Рахман и Абдул Кадер Мункайла Хамани и Айванг Дуан, 2021 год. « Совместное влияние планирования орошения и режима азотных удобрений на рост, урожайность и эффективность использования воды при капельном орошении озимой пшеницы », Устойчивое развитие, MDPI, Open Access Journal, vol. 13 (5), страницы 1-17, март.
- Gardenas, A.I. И Hopmans, J.W. И Хэнсон, Б.И Симунек Дж., 2005. « Двумерное моделирование выщелачивания нитратов для различных сценариев фертигации при микроорошении ,» Управление водными ресурсами в сельском хозяйстве, Elsevier, vol. 74 (3), страницы 219-242, июнь.
- Сампаткумар, Т., Пандиан, Б.Дж. и Рангасвами, М.В. И Маникасундарам П. и Джеякумар П., 2013. « Влияние дефицитного орошения на рост, урожайность и параметры урожайности в последовательности посевов хлопок – кукуруза ,» Управление водными ресурсами в сельском хозяйстве, Elsevier, vol.130 (C), страницы 90-102.
- Арбат, Г. и Росельо, А., Доминго Оливе, Ф. и Пуч-Баргес, Х. и Гонсалес Ллинас, Э. и Дуран-Рос, М., Пухоль, Х. и Рамирес де Картахена, Ф., 2013. « Распределение почвенной воды и нитратов при капельном орошении кукурузы, получающей навоз свиней ,» Управление водными ресурсами в сельском хозяйстве, Elsevier, vol. 120 (C), страницы 11-22.
- Лю, Хайцзюнь и Ван, Сюмин и Чжан, Сиань и Чжан, Ливэй и Ли, Ян и Хуанг, Гуаньхуа, 2017.« Оценка реакции роста, урожайности и эффективности использования воды кукурузы (Zea mays L.) на капельное орошение в условиях мульчи в ирригационном районе Хетао в Китае », Управление водными ресурсами в сельском хозяйстве, Elsevier, vol. 179 (C), страницы 144-157.
- Эль-Хендави, Салах Э. и Котаб, Махер А. и Аль-Сухайбани, Насер А. и Шмидхальтер, Урс, 2014. « Оптимальные сочетания между скоростью полива и уровнями глицинбетаина для повышения урожайности и эффективности использования воды капельным орошением кукурузы, выращиваемой в засушливых условиях », Управление водными ресурсами в сельском хозяйстве, Elsevier, vol.140 (C), страницы 69-78.
- Lekakis, E.H. И Георгиу, П. & Павлатоу-Ве, А. & Антонопулос, В.З., 2011. « Влияние фиксированного частичного осушения корневой зоны и текстуры почвы на динамику воды и растворенных веществ в известковых почвах и урожай кукурузы », Управление водными ресурсами в сельском хозяйстве, Elsevier, vol. 101 (1), страницы 71-80.
- Аль-Огайди, Ахмед А.М. И Ваяйок, Эмрун и Роушон, М.К. И Абдулла, Ахмад Фикри, 2017. « Влияние намагниченной воды на динамику воды в почве в системах капельного орошения ,» Управление водными ресурсами в сельском хозяйстве, Elsevier, vol.180 (PA), страницы 70-77.
- Chehab, Hechmi & Tekaya, Mariem & Mechri, Beligh & Jemai, Abdelmajid & Guiaa, Mohamed & Mahjoub, Zoubeir & Boujnah, Dalenda & Laamari, Salwa & Chihaoui, Badreddine & Zakhama, Houda & Hammami, 2017. « Влияние супервпитывающего полимера Stockosorb® на давление тургора листьев, характеристики деревьев и качество масла оливковых деревьев сорта Chemlali, выращенных в полевых условиях в засушливом регионе Туниса », Управление водными ресурсами в сельском хозяйстве, Elsevier, vol.192 (C), страницы 221-231.
- Андреу, Л. и Хопманс, Дж. У. и Шванкль, Л. Дж., 1997. « Пространственное и временное распределение водного баланса почвы для миндального дерева с капельным орошением ,» Управление водными ресурсами в сельском хозяйстве, Elsevier, vol. 35 (1-2), страницы 123-146, декабрь.
- Мубарак, Ибрагим и Майлхол, Жан Клод и Ангуло-Харамильо, Рафаэль и Буарфа, Сами и Руэль, Пьер, 2009. « Влияние временной изменчивости гидравлических свойств почвы на моделирование переноса воды при высокочастотном капельном орошении ,» Управление водными ресурсами в сельском хозяйстве, Elsevier, vol.96 (11), страницы 1547-1559, ноябрь.
- Мартинес-Коб, А. и Фаси, Дж. М., 2010. « Эвапотранспирация из оливкового сада после обрезки живой изгороди в полузасушливом районе северо-восточной Испании ,» Управление водными ресурсами в сельском хозяйстве, Elsevier, vol. 97 (3), страницы 410-418, март.
- Чаухдари, Джунаид Наваз и Бахш, Аллах и Энгель, Бернард А. и Рагаб, Рагаб, 2019. « Улучшение производства кукурузы за счет внедрения эффективных методов фертигации: экспериментальный подход и моделирование », Управление водными ресурсами в сельском хозяйстве, Elsevier, vol.221 (C), страницы 449-461.
- Робель Адмасу и Авраам В. Майкл и Тилахун Хордофа, 2019. « Старший научный сотрудник ирригации, Центр сельскохозяйственных исследований Мелкасса, Эфиопия ,» Международный журнал наук об окружающей среде и природных ресурсов, Juniper Publishers Inc., vol. 16 (4), страницы 83-87, январь.
- Кылыч, Мурат, 2020. « Новый аналитический метод для оценки трехмерной объемной картины увлажнения в системе капельного орошения », Управление водными ресурсами в сельском хозяйстве, Elsevier, vol.228 (С).
Исправления
Все материалы на этом сайте предоставлены соответствующими издателями и авторами. Вы можете помочь исправить ошибки и упущения. При запросе исправления укажите идентификатор этого элемента: RePEc: eee: agiwat: v: 184: y: 2017: i: c: p: 114-123 . См. Общую информацию о том, как исправить материал в RePEc.
По техническим вопросам, касающимся этого элемента, или для исправления его авторов, заголовка, аннотации, библиографической информации или информации для загрузки, обращайтесь:.Общие контактные данные провайдера: http://www.elsevier.com/locate/agwat .
Если вы создали этот элемент и еще не зарегистрированы в RePEc, мы рекомендуем вам сделать это здесь. Это позволяет привязать ваш профиль к этому элементу. Это также позволяет вам принимать потенциальные ссылки на этот элемент, в отношении которых мы не уверены.
Если CitEc распознал библиографическую ссылку, но не связал с ней элемент в RePEc, вы можете помочь с этой формой .
Если вам известно об отсутствующих элементах, цитирующих этот элемент, вы можете помочь нам создать эти ссылки, добавив соответствующие ссылки таким же образом, как указано выше, для каждого ссылочного элемента. Если вы являетесь зарегистрированным автором этого элемента, вы также можете проверить вкладку «Цитаты» в своем профиле RePEc Author Service, так как там могут быть некоторые цитаты, ожидающие подтверждения.
По техническим вопросам, касающимся этого элемента, или для исправления его авторов, заголовка, аннотации, библиографической информации или информации для загрузки, обращайтесь: Кэтрин Лю (адрес электронной почты указан ниже).
Сокращения: