Как расшифровывается допог: «ДОПОГ как расшифровывается?» — Яндекс.Кью
ДОПОГ. Что нужно знать про международную перевозку опасных грузов – Дзен – АТИ, Центр: Система грузоперевозок
Чтобы не потеряться в обширных правилах перевозки опасных грузов и иметь в запасе необходимое количество подсказок, предлагаем ознакомиться с этой статьей.
Благодаря ей можно более подробно понять структуру положения ДОПОГ, его расшифровку, определения и получить некоторые практические рекомендации для правильного оформления документа.
Что такое ДОПОГ
Аббревиатура ДОПОГ расшифровывается как дорожная перевозка опасных грузов. Это международный договор, который был подписан в Женеве еще полвека назад. Главная задача данного соглашения заключается в объединении правил по международной дорожной перевозке опасных грузов по территории стран ЕС с использованием специального автотранспорта.
Определение «опасного груза»
Зачастую под опасным грузом подразумевают разновидности отходов, но это могут быть, по сути, любые вещества, которые при повреждении целостности упаковки и «утечке» могут нанести вред природе, людям и животным.
Основные положения соглашения для перевозчиков опасных грузов освещают темы:
- допустимой тары для перевозки груза, исходя из его класса;
- маркировки емкостей;
- правил для автотранспорта при транспортировке груза.
Оформление ДОПОГ на транспортировку
Чтобы иметь возможность перевозить опасные грузы по РФ и странам ЕС, которые подписали соглашение, шофер должен обязательно получить свидетельство ДОПОГ. Все подобные грузы делятся на девять основных категорий. Шофер должен получить разрешения на конкретные категории, по которым он прошел обучение. В свидетельстве в двух графах отмечаются классы грузов.
Для получения очередной категории необходимо снова проходить обучение и сдавать экзамены. Сопроводителю груза или экспедитору тоже потребуются свидетельства.
Получить такие свидетельства можно на специальных курсах, утвержденных Минтрансом РФ. Их длительность составляет пять дней. Существует основной курс, куда входят базовые понятия и правила по технике безопасности, особенности поведения водителей и т. д.
д.
Свидетельство будет выдано только после прохождения всех курсов и успешной сдачи экзаменов. К экзаменам допускаются только специалисты и водители, которые полностью прошли курсы в течение пяти дней. Раньше было достаточно одной лекции, но правила изменились после 2013 года. Для водителей цистерн есть отдельные курсы, где им будут разъяснены специфические моменты транспортировки, правила заливки/опорожнения и т.д. Также есть курсы по перевозке радиоактивных и взрывчатых веществ.
Получить свидетельство можно только на пять лет. Оно оформляется на двух языках, один из которых обязательно русский, а второй — иностранный: как правило, английский (хотя допустимы также немецкий или французский языки).
Как опасным веществам и грузам присваивается категория?
При перевозке специфических веществ/грузов необходимо определить их разновидность и оценить уровень опасности. Особенно это касается взрывчатых изделий (АDR).
В настоящее время опасные грузы делятся на девять классов. Для некоторых веществ и изделий разработаны подклассы:
Для некоторых веществ и изделий разработаны подклассы:
- Изделия, подверженные взрыву или самовозгоранию.
- Газовые вещества различных видов: способные воспламеняться, сжaтые газы, растворяющиеся под давлением.
- Легковозгораемые вещества. Жидкости, содержащие твердые материалы, способные выделять самовозгораемые пары.
- Легковозгораемые изделия, способные загореться во время перевозки из-за влияния внешних факторов: соприкосновения с другими грузами, контакта с влагой, нагрева и так далее.
- Изделия с повышенным уровнем радиации.
- Вещества, подверженные окислению. Кислородовыделяемые, воспламеняющиеся пероксиды.
- Повреждающие химические вещества, грозящие кожными ожогами, проникновением в слизистые, дыхательные органы и т.д.
- Вещества, выделяющие яды, приводящие к летальному исходу, сильной интоксикации и поражениям кожи, слизистой.
- Вещества, не представляющие большой опасности при перевозке. Имеют определенный перечень требований к хранению и транспортировке.


Подробная классификация позволяет определить оптимальную емкость для его перевозки. При перевозке опасного груза его класс указывается в товарно-сопроводительных документах. Также прозрачная и понятная классификация упрощает проведение инструктажа для перевозчиков.
Особенности грузоперевозки в России: подводные камни
Для свободного передвижения автомобиля с опасным грузом необходимо наличие документа — разрешения ДOПOГ/АDR.
Местное представительство Российского транспортного надзора отвечает за оформление указанных документов для выезда за границу территории или движения по федеральным трассам. В компетенции региональных управлений государственного автодорожного надзора лишь выдача разрешений для передвижения по местным дорогам на одной региональной территории.
Заявка на получение документа оформляется на стандартном бланке.
К заявке прикрепляется:
- ДOПOГ-свидетельство на транспортное средство и водителя.
- Аварийная/информационная карта.
- Маршрут транспортного средства.

При составлении маршрута важно помнить про:
- Запрет на движение спецтранспорта через зоны отдыха, через природные заказники, вблизи объектов культурного наследия и вблизи крупных заводов.
- Необходимость проставлять метки для участков, безопасных для остановки, стоянок и заправок.
Органы ГИБДД должны быть осведомлены, если транспортировка осуществляется на нескольких автомобилях. В таком случае машина, находящаяся впереди, отмечается маяком оранжевого или желтого цвета.
Согласно приказу от 2012 года, на автомобили, выбранные для перевозки, устанавливается спутниковая система ГЛОНАCC.
ДОПОГ — маркировка опасного багажа
Токсичные вещества упаковываются по-разному. Газовые или жидкие субстанции помещаются в баллоны и цистерны. Оптимальная емкость для твердых материалов — ящики, контейнеры, коробы. Порошковые вещества укладываются в мешки.
Наличие пометки — обязательное условие транспортировки. Как правило, маркировка имеет квадратную форму, пиктограмму и символику с обозначением категории. Допустимые параметры — не меньше 10 см по одной стороне.
Как правило, маркировка имеет квадратную форму, пиктограмму и символику с обозначением категории. Допустимые параметры — не меньше 10 см по одной стороне.
Маркировка должна отвечать требованиям ГОСТ-стандарта. В противном случае перевозчики рискуют получить штраф.
Кто имеет право на получение свидетельства ДОПОГ
Согласно приказу Министерства транспорта России под номером 282, начиная с 1 января 2021 года, для транспортировки опасных грузов допускаются шоферы, которые обладают:
- водительскими правами РФ соответствующей категории;
- свидетельством ДОПОГ.
Помимо этого к шоферу предъявляются следующие требования:
- Исходя из вида автотранспорта, помимо основных знаний по вождению, полученных на курсах автошколы, дополнительно шофер должен знать и общие правила, описанные в соглашении ДОПОГ по перевозке опасных грузов.
- Исходя из вида управляемого автотранспорта, помимо основных знаний по предоставлению первой медицинской помощи, полученных на курсах автошколы, шофер должен обладать и другими знаниями по оказанию первой помощи в случае с происшествием во время перевозки опасного груза. Также водитель должен следить за загрузкой/погрузкой опасного груза, в случае необходимости выполнять действия по ликвидации или локализации последствий аварии автомобиля, который транспортирует опасный груз.
- Чтобы шофер был допущен к управлению автотранспортом, который выполняет транспортировку опасного груза, он должен иметь минимум трехгодичный водительский стаж. Помимо всего прочего, он в течение последнего года не должен привлекаться к наказаниям в виде лишения прав или административного ареста за нарушение правил дорожного движения.
 Также водитель должен следить за загрузкой/погрузкой опасного груза, в случае необходимости выполнять действия по ликвидации или локализации последствий аварии автомобиля, который транспортирует опасный груз.
Также водитель должен следить за загрузкой/погрузкой опасного груза, в случае необходимости выполнять действия по ликвидации или локализации последствий аварии автомобиля, который транспортирует опасный груз.Требования соглашения ДОПОГ
Эти правила представляют собой одновременно и подсказку по правильному оформлению опасного груза, подготовке подходящего транспорта и пути его маршрута. Данные правила были приняты Россией еще в 1995 году, но уже в 1999 были внесены новые поправки. Среди главных пунктов соглашения можно отметить:
- Основные положения.
- План транспортировки.
- Работа с транспортными компаниями клиентов.
- Техническое обеспечение транспортировки грузов.
- Требования к водителям и сопровождающим груз.
- Дополнительные требования к некоторым классам опасных грузов.
- Точная классификация подобных видов груза.
- Список и подробное описание подобных грузов.
- Образцы информационных документов в виде графиков и таблиц.
- Другие приложения относительно правильности оформления документов, упаковки и т.д.

Что касается алгоритма планирования транспортировки, он выглядит следующим образом:
- сначала необходимо получить все необходимые разрешения и сертификаты на перевозку опасного груза вне границ России;
- получить разрешение на перевозку очень опасных грузов;
- заключить контракт на транспортировку;
- подготовить водителя, экспедитора и/или сопровождающего груз;
- обозначить путь на карте;
- принять груз;
- сформировать систему оповещения об опасной транспортировке;
- выполнить погрузку/разгрузку;
- соблюдать правила движения средств;
- соблюдать нюансы при перевозке грузов разных классов в одном автотранспорте;
- правильно обращаться с использованной упаковкой;
- освоить механизм действий во время аварий.

Есть еще один вид перевозки опасного груза посредством железнодорожного транспорта. Тем, кто занимается железнодорожными перевозками товаров такой категории, необходимо знать правила и требования по их транспортировке.
Необходимо понимать, что правила ДОПОГ должны в обязательном порядке быть изучены компаниями, которые занимаются перевозками, но и владелец опасного груза тоже должен их знать. В противном случае могут возникнуть проблемы, которые повлекут не только финансовые потери.
Комплекты ADR. Оборудование ADR
В данной категории представлены комплекты ADR и сопуствующее оборудование ADR
 Действует договор на территории всех стран Евросоюза, а также в Казахстане, Азербайджане и Марокко. В России аналогичный ему документ называется ДОПОГ.
Действует договор на территории всех стран Евросоюза, а также в Казахстане, Азербайджане и Марокко. В России аналогичный ему документ называется ДОПОГ. Комплект ADR – это комплект дополнительного оборудования и средств индивидуальной защиты для транспортного средства, перевозящего опасный груз и для его членов экипажа. Набор ADR включает все предметы, которые должны быть в обязательном порядке на транспортном средстве при перевозке опасного груза, согласно требованиям ДОПОГ/ADR, за исключением огнетушителей.
Все ADR комплекты различаются по количеству и наименованию составляющих и комплектуются соответственно классу опасности перевозимого груза, либо по требованиям клиентов. Автомобиль, перевозящий опасные грузы, в соответствии с требованиями ДОПОГ 2013, обязательно должен быть оборудован следующим необходимым снаряжением, оборудованием и материалами:
- Противооткатный башмак,
- Вертикально (самостоятельно) стоящих предупреждающих аварийных знака -2 шт,
- Лопата, пластиковые мешки, пластиковый контейнер для сбора абсорбента, дренажная ловушка (касается грузов класса 3, 4. 1, 4.3, 8, 9)
- Абсорбент (касается грузов класса 3)
- Огнетушитель 2 кг переносной
- Огнетушитель не менее 6 кг на каждую транспортную единицу в легкодоступном месте
- Оранжевые информационные таблицы, обозначающие опасный груз, крепящиеся на транспортные средства
 1, 4.3, 8, 9)
1, 4.3, 8, 9)Комплект личной защиты экипажа на каждого члена экипажа должен состоять из:
- Предупреждающий (сигнальный) жилет
- Фонарь (характеристики согласно ДОПОГ)
- Защитные перчатки
- Защитные очки
- Защитная маска (касается грузов класса 2.3 и 6.1)
- Жидкость для полоскания глаз (не касается грузов класса 1, 1.4, 1.5, 1.6, 2.1, 2.2, 2.3)
Все оборудвоание ADR подбираются (согласно ДОПОГ) соответственно классу опасности (ромбу) перевозимого вещества. В случае присвоения веществу нескольких классов опасности, основанием для выбора ADR комплекта является первый (основной) знак класса опасности (ромб) указанный в таблицах ДОПОГ (текст ДОПОГ). Транспортные средства перевозящие опасные грузы в упаковках и в освобожденных количествах ADR комплектами не комплектуются.
Транспортные средства перевозящие опасные грузы в упаковках и в освобожденных количествах ADR комплектами не комплектуются.
В нашем магазине представлен широкий ассортимент комплектов ADR и сопусвтующего оборудования ADR, которое удовлетворит даже самого взыскательного клиента. Вы можете сформировать заказ самостоятельно или с помощью наших квалифицированных менеджеров, которые проконсультируют Вас по любому интересующему вопросу о наборах ADR и оборудовании ADR.
ДОПОГ — ГК «ЮГРАНАВИТЕХ»
Дорожная Перевозка Опасных Грузов автотранспортом
Что такое ДОПОГ?
В соответствии с требованиями технического регламента таможенного союза (ТР ТС 018/2011) автомобиль, предназначенный для перевозки опасных грузов должен отвечать требованиям Правил ЕЭК ООН № 105 (Единообразные предписания, касающиеся официального утверждения транспортных средств, предназначенных для перевозки опасных грузов, в отношении их конструктивных особенностей), которые практически дублируют требования Главы 9.
Ранее существовали чисто российские требования по перевозкам этой области (ПОГАТ), однако сейчас Российская Федерация приняла в полном объеме европейские требования (ДОПОГ).
ДОПОГ дословно расшифровывается как – Дорожная Перевозка Опасного Груза (или грузов, если их несколько видов).
ДОПОГ представляет собой соглашение между государствами и не предусматривает наличие какого-либо общего органа для обеспечения соблюдения его положений. На практике проверки на автомагистралях осуществляются Договаривающимися сторонами и несоблюдение положений Соглашения может привести к возбуждению национальными органами иска против нарушителей в соответствии с их внутригосударственным законодательством.
Требования ДОПОГ по оснащению автотранспорта
Согласно ДОПОГ предусмотрены следующие требования к переоборудованию автомобилей для перевозки опасных грузов:
1. Требования к электропроводке
— Защита проводки от механических и термических повреждений.
Электрическая проводка автомобиля должна быть защищена от механических повреждений, от перегрева и воспламенения, крепление проводки должно быть прочным и надежным. п. 9.2.2.2.2 ДОПОГ. Данное требование распространяется на транспортные средства типа: EX/III, AT, FL, OX.
— Наличие дистанционного выключателя массы.
В соответствии с требованием ЕЭК ООН №105 транспортное средство должно быть оборудовано устройством дистанционным выключателем массы.
— Наличие главного выключателя аккумуляторной батареи (выключатель массы)
В соответствии с п. 9.2.2.8 ДОПОГ в кабине автомобиля должна находиться кнопка выключателя массы для быстрого разрыва электрических цепей.
2. Требования к тормозному оборудованию
— Наличие антиблокировочной системы торможения (АБС).
В связи с вступлением РФ в Таможенный Союз и вступлением новой редакции ДОПОГ от 01.01.2015 г. в обязательном порядке необходимо наличие антиблокировочной системы торможения (ABS) на всех автомобилях, осуществляющих перевозку опасного груза.
Согласно п. 9.2.3.1.1 ДОПОГ, которое ссылается на Правила ЕЭК ООН №13, необходимо выполнение требований к тормозной системе автомобиля. Данное требование говорит об обязательной установке Антиблокировочной Системы Торможения (АБС).
— Регистрация изменений конструкции ТС.
Необходимо оформление документов и регистрации внесения изменений в конструкцию транспортного средства.
3. Требования к топливному баку
— Защита топливного бака.
Согласно пункта 9.2.4.3 ДОПОГ в котором сказано о требованиях к монтажу защиты топливного бака автомобиля. Топливный бак должен быть расположен таким образом, что при вытекании топлива на землю, оно не попадало на нагретые агрегаты транспортного средства. На горловине топливного бака должна присутствовать крепкая и герметичная крышка.
4. Требования к системе выпуска выхлопных газов
— Защита груза от перегрева.
Система выпуска выхлопных газов (включая выхлопные трубы) должна быть расположена или защищена таким образом, чтобы груз не подвергался никакой опасности перегрева или воспламенения.
5. Требования к топливным обогревательным приборам
— Установка согласно правил ЕЭК ООН №122.
Установка воздушных и жидкостных отопителей, сепараторов по правилам Европейская экономическая комиссия ООН №122 «Единообразные технические предписания, касающиеся официального утверждения транспортных средств категорий M, N и O в отношении их систем отопления».
— Защита груза от перегрева выхлопной трубой.
Выхлопная система воздушных и жидкостных отопителей должна быть смонтированная, защищена и расположена в местах, где груз не смог бы быть подвергнут опасности и возгоранию.
6. Наличие устройства ограничения скорости
— Установка устройства ограничения скорости (УОС).
Требования Европейской Экономической Комиссии ООН №89 и согласно ДОПОГ п. 9.2.5 обязуют автомобили оснащать или иметь в заводской комплектации устройства ограничения скорости (УОС). Всем транспортным средствам максимальной массой более 3,5 тонн в обязательном порядке необходимо иметь УОС в соответствии с техническими требованиями Правил ЕЭК ООН № 89 с поправками.
7. Наличие тахографа
— Наличие тахографа.
В соответствии с приказом №36 Министерства транспорта Российской Федерации и требованиями Технического Регламента Таможенного Союза (ТР ТС) транспортное средство должно быть оснащено тахографом.
— Правильная эксплуатация тахографа.
Тахограф должен быть установлен по правилам тахографического контроля, активирован и вовремя откалиброван. У водителей должны быть личные карты водителя.
8. Наличие системы ГЛОНАСС
— Наличие спутниковой навигации ГЛОНАСС или ГЛОНАСС/GPS.
В соответствии с Техническим регламентом о безопасности колесных транспортных средств, на транспортных средствах, осуществляющих перевозку опасных грузов и пассажиров, численностью свыше 9 человек, необходимо наличие оборудования спутниковой навигации ГЛОНАСС или ГЛОНАСС/GPS.
9. Требование к спецсигналам
— Обязательное наличие проблескового маячка.
Обязательное наличие маяка. Проблесковый маяк должен быть включен за весь период движения транспортного средства. Рекомендуем установить два маяка, на случай выхода из строя одного.
Рекомендуем установить два маяка, на случай выхода из строя одного.
— Обязательное наличие контурной маркировки.
Обязательное нанесение контурной маркировки на транспортное средство.
Переоборудование транспортных средств по ДОПОГ
В Группе Компаний ЮГРАНАВИТЕХ вы можете получить полный комплекс услуг по переоборудованию транспортных средств, перевозящих опасные грузы согласно требованиям ДОПОГ и техрегламента.
Услуги включают в себя установку:
— главного выключателя аккумуляторной батареи (выключатель массы)
— устройства ограничения скорости
— тахографа
— системы мониторинга GPS/ГЛОНАСС
— проблесковых маячков
— и другого дополнительного оборудования
Отметим, что внесения изменений в конструкцию транспортного средства требуют обязательной регистрации. Специалисты нашей компании проведут все необходимые работы по узаконению изменений в конструкции ТС.
Для получения подробной консультации по вопросам обучения или оснащения транспорта, вы можете обратиться к нашим специалистам по телефону +7 912 080 2696 +7 951 418 9949
Мы поможем Вам определиться с выбором необходимого оборудования и проведем монтажные работы любого уровня сложности.
Что позволяет нам утверждать, что мы — лучшие? — Компания ПоГАТ
Еще великий маг Воланд в булгаковской «Мастер и Маргарите» утверждал, что «факты – самая упрямая в мире вещь». Мы – не волшебники, но полностью с ним согласны.
Взгляните и Вы на них:
Во-первых, весь используемый транспорт – наша собственность. Он многократно прошел проверки на надежность. Именно поэтому мы не зависим от недостатков и проблем сторонних компаний-экспедиторов. Вдобавок наша ответственность и все наши машины застрахованы.
Во-вторых, мы постоянно контролируем весь процесс транспортировки: от погрузки до прибытия автомобиля к грузополучателю. Мы знаем, что происходит с нашим транспортом.
В-третьих, в своей работе мы руководствуемся принципами, на которых строится сильная современная армия. Мы не понаслышке знаем, что такое дисциплина и порядок. Поэтому мы абсолютно не приемлем хаоса и неопределенности. Мы делаем ответственное дело и здесь не место случайностям.
В-четвертых, мы предлагаем свои партнерам машины, способные перевозить от 500 кг до 20 тонн опасных и особо опасных веществ. Наша деятельность универсальна – мы поможем и мелкому и крупному бизнесу в решении его проблем.
Наша деятельность универсальна – мы поможем и мелкому и крупному бизнесу в решении его проблем.
В-пятых, перечень перевозимых нами опасных веществ включает в себя все классы.
В-шестых, каждый наш водитель прошел специальное обучение и имеет ДОПОГ-свидетельства. Что это такое? ДОПОГ расшифровывается как ДОрожные Перевозки Опасных Грузов. Без подобного документа наши перевозки были бы такими же незаконными, как вождение без водительского удостоверения. Мы знаем и соблюдаем закон.
В-седьмых, перед тем, как заключить контракт, мы проверяем соответствие заявленного к перевозке груза российским и международным стандартам перевозки опасных и особо опасных веществ. Только после этого мы выпускаем машину в рейс. Наш принцип: Никаких компромиссов в ущерб Качеству! Попытки «договориться» в обход Закона ведут к расторжению договора и внесения данных клиента в «черный список».
В-восьмых, мы не только перевозим опасные грузы и отходы. Мы занимаемся всем комплексом работ сразу: и перевозкой, и подготовкой документов, и вооруженной охраной на всем пути следования.
В-девятых, как транспортная компания мы имеем очень удобное расположение: всего сотня метров от развязки с Московской Кольцевой Автомобильной Дорогой. Это позволяет нам экономить время и деньги как свои, так и клиентов.
В-десятых, наш подвижной состав состоит только из качественных автомобилей иностранного производства, количество которых постоянно увеличивается. Все автомобили нашей компании имеют Свидетельства о допуске транспортных средств к перевозке всех опасных грузов.
В-одиннадцатых, в нашем автопарке всегда имеется дежурное транспортное средство, готовое в любое время выехать к клиенту или заменить вышедший из строя автомобиль. Также в офисе действует круглосуточная телефонная диспетчерская служба.
И наконец, у нас общие с нашими клиентами ценности – высочайшее качество, стопроцентная надежность, нацеленность на абсолютный результат и постоянное стремление к дальнейшему развитию.
Разгадка загадок приручения собак
Zool Res. 2020 18 марта; 41(2): 97–104.
Чжэ Чжан
1 Ключевая государственная лаборатория сохранения и использования биоресурсов в Юньнани, Школа наук о жизни, Юньнаньский университет, Куньмин, Юньнань 650091, Китай,
Сабер Хедерзаде
2 Государственная ключевая лаборатория генетических ресурсов и эволюции, Банк зародышевой плазмы диких видов, Куньминский институт зоологии, Китайская академия наук, Куньмин, Юньнань 650223, Китай,
3 Куньминский колледж наук о жизни Университета Китайской академии наук, Куньмин, Юньнань 650204, Китай,
Ян Ли
1 Ключевая государственная лаборатория сохранения и использования биоресурсов в Юньнани, Школа наук о жизни, Юньнаньский университет, Куньмин, Юньнань 650091, Китай,
1 Ключевая государственная лаборатория сохранения и использования биоресурсов в Юньнани, Школа наук о жизни, Юньнаньский университет, Куньмин, Юньнань 650091, Китай,
2 Государственная ключевая лаборатория генетических ресурсов и эволюции, Банк зародышевой плазмы диких видов, Куньминский институт зоологии, Китайская академия наук, Куньмин, Юньнань 650223, Китай,
3 Куньминский колледж наук о жизни Университета Китайской академии наук, Куньмин, Юньнань 650204, Китай,
Поступила в редакцию 5 июля 2019 г . ; Принято 6 января 2020 г.
; Принято 6 января 2020 г.
Abstract
Домашняя собака, как очень успешная модель одомашнивания, хорошо известна как любимый компаньон человека.Изучение истории его одомашнивания должно пролить свет на наше понимание доисторического развития человеческой культуры и продуктивности. Кроме того, изучение механизмов, лежащих в основе морфологических и поведенческих признаков, связанных с синдромом одомашнивания псовых, имеет значение не только для научных исследований, но и для медицинских исследований человека. Текущая разработка междисциплинарной базы данных генома собак, которая включает в себя огромные данные по омике, существенно улучшила наше понимание генетического состава собак. Здесь мы рассмотрели недавние достижения, связанные с оригинальной историей и генетической основой, лежащей в основе адаптации к окружающей среде и фенотипического разнообразия у домашних собак, которые должны обеспечить перспективы улучшения коммуникативных отношений между собаками и людьми.
Здесь мы рассмотрели недавние достижения, связанные с оригинальной историей и генетической основой, лежащей в основе адаптации к окружающей среде и фенотипического разнообразия у домашних собак, которые должны обеспечить перспективы улучшения коммуникативных отношений между собаками и людьми.
Ключевые слова: Собака, История приручения, Адаптация, Фенотипическое разнообразие, Внутреннее наследие
ВВЕДЕНИЕ
Собаки, как первое одомашненное млекопитающее, взаимодействовали с людьми на протяжении тысячелетий (Freedman & Wayne, 2017).Это одомашнивание происходило в два основных этапа: первоначальное приручение дикого серого волка до первичной собаки и последующее улучшение этих местных собак до различных современных пород. На первом этапе весьма специфическое поведение, ценимое кочевым обществом охотников-собирателей, такое как выслеживание и поедание добычи, определяло первоначальную эволюцию первичных собак в начале одомашнивания (Ostrander et al. , 2017). Во время этого процесса отбор был не единственным механизмом эволюции, поскольку генетический дрейф, вероятно, также приводил к тому, что подходящие, непригодные или нейтральные аллели фиксировались из-за узких мест или эффектов основателя (Doebley, 1989; Eyre-Walker et al., 1998). Более того, признаки, обычно связанные с одомашниванием, не отбирались напрямую, а скорее представляли собой побочные эффекты ослабления отбора (Dobney & Larson, 2006). Например, одомашнивание ослабило селективное давление на самцов собак, чтобы они соперничали за партнеров. Таким образом, гены, участвующие в производстве признаков, имеющих отношение к этому начинанию, больше не подвергались селективному давлению и, таким образом, стали фактически нейтральными или даже вредными, поскольку они включали отвлечение ресурсов от признаков, подвергающихся отбору, таких как прирученность.С появлением сельскохозяйственных обществ собаки приспособились к введению крахмала в свой рацион, что стало важным шагом в их эволюции от волка (Арендт и др.
, 2017). Во время этого процесса отбор был не единственным механизмом эволюции, поскольку генетический дрейф, вероятно, также приводил к тому, что подходящие, непригодные или нейтральные аллели фиксировались из-за узких мест или эффектов основателя (Doebley, 1989; Eyre-Walker et al., 1998). Более того, признаки, обычно связанные с одомашниванием, не отбирались напрямую, а скорее представляли собой побочные эффекты ослабления отбора (Dobney & Larson, 2006). Например, одомашнивание ослабило селективное давление на самцов собак, чтобы они соперничали за партнеров. Таким образом, гены, участвующие в производстве признаков, имеющих отношение к этому начинанию, больше не подвергались селективному давлению и, таким образом, стали фактически нейтральными или даже вредными, поскольку они включали отвлечение ресурсов от признаков, подвергающихся отбору, таких как прирученность.С появлением сельскохозяйственных обществ собаки приспособились к введению крахмала в свой рацион, что стало важным шагом в их эволюции от волка (Арендт и др. , 2016). За последние несколько сотен лет фенотипическая радиация среди одомашненных собак ускорила процесс разведения, что привело к развитию почти 400 пород, выполняющих специализированные функции в человеческом обществе, с четкими морфологическими и поведенческими различиями, такими как большие или маленькие, худощавые или приземистые, и независимые или лояльные (Freedman & Wayne, 2017).Это улучшение селекции было в значительной степени обусловлено экстремальным влиянием искусственного отбора с резким увеличением вредных аллелей, т. е. «внутренних издержек» (Cruz et al., 2008). Огромное разнообразие в морфологии и поведении современных пород подчеркивает силу и привлекательность искусственного отбора, который широко использовался в течение длительного периода и обладает неоценимым потенциалом.
, 2016). За последние несколько сотен лет фенотипическая радиация среди одомашненных собак ускорила процесс разведения, что привело к развитию почти 400 пород, выполняющих специализированные функции в человеческом обществе, с четкими морфологическими и поведенческими различиями, такими как большие или маленькие, худощавые или приземистые, и независимые или лояльные (Freedman & Wayne, 2017).Это улучшение селекции было в значительной степени обусловлено экстремальным влиянием искусственного отбора с резким увеличением вредных аллелей, т. е. «внутренних издержек» (Cruz et al., 2008). Огромное разнообразие в морфологии и поведении современных пород подчеркивает силу и привлекательность искусственного отбора, который широко использовался в течение длительного периода и обладает неоценимым потенциалом.
Домашние собаки распространились по всему миру после миграции людей и, таким образом, адаптировались к очень разным условиям.Это долгосрочное сотрудничество не только способствовало гораздо более тесным отношениям между людьми и собаками, чем с любым другим домашним животным, но также и схожей генетике, лежащей в основе до 360 болезней, от которых страдают и те, и другие, особенно психические заболевания (например, собачье компульсивное расстройство, болезнь Альцгеймера). , эпилепсия) (De Risio et al., 2015; Dewey et al., 2019; Ostrander et al., 2017; Shearin & Ostrander, 2010b), что может частично быть результатом того же природного давления окружающей среды и аналогичного социального воздействия.Поэтому собаки являются хорошими моделями как для эволюционных, так и для патологических исследований.
, эпилепсия) (De Risio et al., 2015; Dewey et al., 2019; Ostrander et al., 2017; Shearin & Ostrander, 2010b), что может частично быть результатом того же природного давления окружающей среды и аналогичного социального воздействия.Поэтому собаки являются хорошими моделями как для эволюционных, так и для патологических исследований.
Объединение молекулярной филогенетики, биоинформатики и биотехнологии помогло разгадать загадки истории одомашнивания собак. Здесь мы обобщили ход одомашнивания собак, включая время и происхождение, адаптацию к различным местным условиям и генетическую основу, лежащую в основе фенотипического разнообразия, чтобы подчеркнуть наше текущее понимание одомашнивания собак и вызвать энтузиазм для дальнейшего разведения в соответствии с генетической информацией.
ПРОИСХОЖДЕНИЕ СОБАК
Археологические данные показывают, что собака была спутником человека со времен мезолита. Данные геномного секвенирования также предполагают, что первоначальная одомашнивание собак предшествовало появлению сельского хозяйства, а вместо этого происходило вместе с охотниками-собирателями (Freedman et al. , 2014). Как генетические, так и фенотипические исследования ясно показали, что собака произошла от вымершей популяции серых волков и была одомашнена в Старом Свете (Fan et al., 2016). Однако продолжаются споры о географическом происхождении монофилогении или полифилогении из-за расхождений в идеях и методах исследования.К счастью, все больше данных проливают свет на происхождение и эволюционные пути собак.
, 2014). Как генетические, так и фенотипические исследования ясно показали, что собака произошла от вымершей популяции серых волков и была одомашнена в Старом Свете (Fan et al., 2016). Однако продолжаются споры о географическом происхождении монофилогении или полифилогении из-за расхождений в идеях и методах исследования.К счастью, все больше данных проливают свет на происхождение и эволюционные пути собак.
Вила и его коллеги исследовали происхождение собак на основе последовательностей контрольной области мтДНК и были первыми, кто предположил полифилию происхождения от множественных предковых популяций волков с событиями обратного скрещивания после первоначального одомашнивания (Vilà et al., 1997). В качестве альтернативы Savolainen et al. (2002) предложил Восточную Азию в качестве единого центра происхождения около 15 000 лет назад из-за наличия богатых генетических и филогеографических вариаций, а также в качестве происхождения австралийских динго (Savolainen et al., 2004).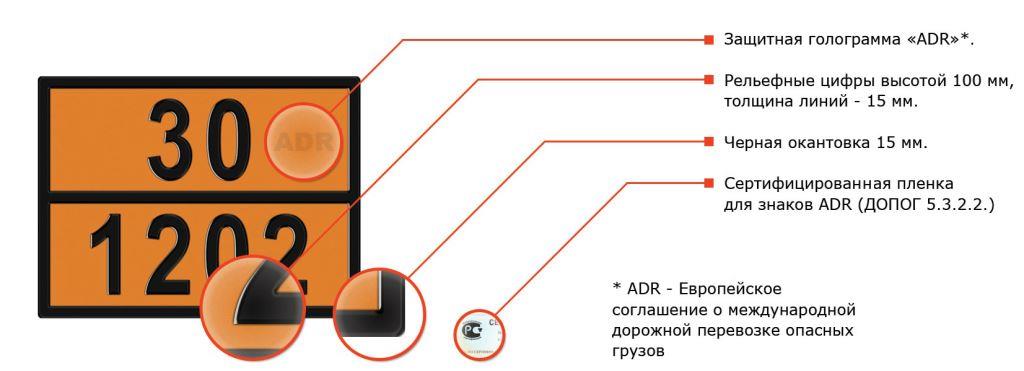 Кроме того, данные также свидетельствуют о том, что индейские собаки произошли от собак Старого Света, которые пересекли Берингов пролив после деятельности человека в позднем плейстоцене (Leonard et al., 2002). Хотя доказательства того, что гаплотипы мтДНК африканских деревенских собак так же разнообразны, как и гаплотипы восточноазиатских, бросают вызов гипотезе восточноазиатского происхождения (Boyko et al., 2009), эта гипотеза подтверждается предыдущими анализами разнообразия митохондриальных и Y-хромосом из почти исчерпывающей выборки. (Динг и др., 2012; Панг и др., 2009).
Кроме того, данные также свидетельствуют о том, что индейские собаки произошли от собак Старого Света, которые пересекли Берингов пролив после деятельности человека в позднем плейстоцене (Leonard et al., 2002). Хотя доказательства того, что гаплотипы мтДНК африканских деревенских собак так же разнообразны, как и гаплотипы восточноазиатских, бросают вызов гипотезе восточноазиатского происхождения (Boyko et al., 2009), эта гипотеза подтверждается предыдущими анализами разнообразия митохондриальных и Y-хромосом из почти исчерпывающей выборки. (Динг и др., 2012; Панг и др., 2009).
Однако гипотеза о восточноазиатском происхождении была опровергнута ядерным геномным полиморфным анализом одиночных нуклеотидов (SNP), который подтверждает среднеазиатское происхождение (Vonholdt et al., 2010). Гипотеза среднеазиатского происхождения основана на двух выводах: (1) породы восточноазиатского происхождения не имеют самого высокого уровня ядерной изменчивости; и (2) большинство пород имеют большинство общих гаплотипов с ближневосточными волками. Тем не менее, этот вывод был опровергнут отсутствием деревенских собак на реке Янцзы, где однородительски унаследованные маркеры представляют наибольшее генетическое разнообразие (Ding et al., 2012; Pang et al., 2009) и значительным смещением SNP, созданных с помощью матричной технологии. Другое исследование, основанное на образцах псовых из Африки, Азии, Европы, Ближнего Востока и Северной Америки, опровергло гипотезу восточноазиатского происхождения с помощью анализа геномного ресеквенирования (Fan et al., 2016). Однако последующее геномное повторное секвенирование образцов псовых на материковой части Евразии выявило миграционный маршрут «из Юго-Восточной Азии», тем самым подчеркнув восточноазиатское происхождение и предшествовавшее началу одомашнивания примерно на 33 000 лет назад (Wang et al., 2016). Исследования древней ДНК также показали, что собака могла быть одомашнена в Восточной и Западной Евразии независимо от различных популяций волков. Затем западно-евразийские собаки были частично заменены восточными собаками после экспансии фермеров ближневосточного неолита в Европу (Frantz et al.
Тем не менее, этот вывод был опровергнут отсутствием деревенских собак на реке Янцзы, где однородительски унаследованные маркеры представляют наибольшее генетическое разнообразие (Ding et al., 2012; Pang et al., 2009) и значительным смещением SNP, созданных с помощью матричной технологии. Другое исследование, основанное на образцах псовых из Африки, Азии, Европы, Ближнего Востока и Северной Америки, опровергло гипотезу восточноазиатского происхождения с помощью анализа геномного ресеквенирования (Fan et al., 2016). Однако последующее геномное повторное секвенирование образцов псовых на материковой части Евразии выявило миграционный маршрут «из Юго-Восточной Азии», тем самым подчеркнув восточноазиатское происхождение и предшествовавшее началу одомашнивания примерно на 33 000 лет назад (Wang et al., 2016). Исследования древней ДНК также показали, что собака могла быть одомашнена в Восточной и Западной Евразии независимо от различных популяций волков. Затем западно-евразийские собаки были частично заменены восточными собаками после экспансии фермеров ближневосточного неолита в Европу (Frantz et al. , 2016; Ollivier et al., 2018). Американские собаки, вероятно, возникли в Восточной Азии и распространились в Америке вместе с людьми (Ni Leathlobhair et al., 2018), но они почти полностью исчезли после прибытия европейцев, оставив самое близкое обнаруживаемое генетическое наследие собачьей трансмиссивной венерической опухоли индивидууму. 000 лет назад.
, 2016; Ollivier et al., 2018). Американские собаки, вероятно, возникли в Восточной Азии и распространились в Америке вместе с людьми (Ni Leathlobhair et al., 2018), но они почти полностью исчезли после прибытия европейцев, оставив самое близкое обнаруживаемое генетическое наследие собачьей трансмиссивной венерической опухоли индивидууму. 000 лет назад.
В совокупности генетический материал, отвечающий различным правилам наследования, использовался для вывода о происхождении собак. Генетические маркеры на митохондриальной ДНК (мтДНК) и Y-хромосомах стабильно наследуются от их материнской линии и отцовской линии соответственно, что облегчает исследование потока генов от материнских и отцовских предков. Однако отсутствие (или почти полное отсутствие) рекомбинации и чувствительность генетического дрейфа делают эти маркеры неспособными расшифровать точную демографическую историю.Полногеномные данные SNP предоставили больше информации, особенно о потоке генов и неполной сортировке клонов (ILS), для филогенетической реконструкции, что привело к несовместимым результатам с результатами, полученными с помощью мтДНК или Y-хромосом для одомашнивания собак (Ding et al. , 2012; Savolainen et al., 2012). al., 2002; Vilà et al., 1997; Vonholdt et al., 2010). К счастью, недавние исследования с использованием различных наследственных маркеров предоставили более убедительные сведения об эволюции диких животных, таких как комары, и домашних животных, таких как собаки и крупный рогатый скот (Frantz et al., 2016; Таворнваттана и др., 2018 г.; Ву и др., 2018). Таким образом, включение различных генетических маркеров в сочетании с крупномасштабными выборками из разных географических регионов и моментов времени, данными глубокого секвенирования и разработкой более продвинутых методологий улучшит наше понимание эволюционного происхождения и генетического состава собаки.
, 2012; Savolainen et al., 2012). al., 2002; Vilà et al., 1997; Vonholdt et al., 2010). К счастью, недавние исследования с использованием различных наследственных маркеров предоставили более убедительные сведения об эволюции диких животных, таких как комары, и домашних животных, таких как собаки и крупный рогатый скот (Frantz et al., 2016; Таворнваттана и др., 2018 г.; Ву и др., 2018). Таким образом, включение различных генетических маркеров в сочетании с крупномасштабными выборками из разных географических регионов и моментов времени, данными глубокого секвенирования и разработкой более продвинутых методологий улучшит наше понимание эволюционного происхождения и генетического состава собаки.
АДАПТАЦИЯ К РАЗЛИЧНЫМ УСЛОВИЯМ
Собака, которая имеет более тесные отношения с человеком, чем любое другое домашнее животное, рассеялась по всему миру вслед за человеком и адаптировалась к различным местным условиям за относительно короткий период времени.Биологи широко рассматривали генетические механизмы, лежащие в основе адаптации. Здесь мы суммируем два основных фактора: изменение рациона питания и адаптацию к окружающей среде.
Здесь мы суммируем два основных фактора: изменение рациона питания и адаптацию к окружающей среде.
Считается, что в процессе превращения волка в собаку решающую роль сыграла диета. В отличие от волков, собаки могут процветать на диете, богатой крахмалом. Десять генов, участвующих в пищеварении и метаболизме жирных кислот, быстро эволюционировали во время одомашнивания собак (Axelsson et al., 2013), что свидетельствует об изменениях в рационе, вызванных одомашниванием.Три гена ( AMY2B , MGAM , SGLT1 ) были идентифицированы как селективные кандидаты, играющие ключевые роли в расщеплении крахмала. Различные породы собак обладают разной способностью перерабатывать крахмал, что может быть связано с вариациями числа копий гена AMY2B , влияющего на активность амилазы (Arendt et al., 2014). Изменения в количестве копий AMY2B у местных собак, современных пород и волков во всем мире показывают, что изменение рациона питания включало в себя вторичный сдвиг, соответствующий развитию и распространению доисторического земледелия в большинстве частей мира, а не единовременно. возникновение во время первоначального одомашнивания (Арендт и др., 2016).
возникновение во время первоначального одомашнивания (Арендт и др., 2016).
Кроме того, адаптация к окружающей среде также привлекла внимание биологов. Тибетский мастиф, местная порода, обитающая на Тибетском нагорье, развила особые фенотипические черты, адаптированные к условиям высокогорья. Анализ данных массива SNP выявил несколько генов с селективными сигнатурами у тибетского мастифа, особенно ген EPAS1 , который, как сообщается, влияет на адаптацию к окружающей среде у жителей Тибета (Li et al., 2014b).Геномное повторное секвенирование собак с разных высот показало, что вариант G305S и несинонимичные мутации в EPAS1 могут снижать сосудистое сопротивление и, таким образом, способствовать транспорту кислорода (Gou et al., 2014). EPAS1 и HBB далее были идентифицированы как кандидаты на конвергентную адаптацию гемоглобина к гипоксии между тибетцами и тибетскими собаками (Wang et al., 2014). С точки зрения Х-хромосомы, которая игнорировалась в предыдущих исследованиях, частота гаплотипа гена AMOT значительно коррелирует с высотой над уровнем моря (Wu et al. , 2016). Более того, полногеномное сканирование тибетского мастифа и тибетского волка указывает на генетическую интрогрессию локусов EPAS1 и HBB , что подразумевает возможность того, что тибетский мастиф быстро приобрел местную адаптацию в результате вторичного контакта со своим диким родственником, тибетским волком, хотя в локусе AMOT таких доказательств не обнаружено (Miao et al., 2017).
, 2016). Более того, полногеномное сканирование тибетского мастифа и тибетского волка указывает на генетическую интрогрессию локусов EPAS1 и HBB , что подразумевает возможность того, что тибетский мастиф быстро приобрел местную адаптацию в результате вторичного контакта со своим диким родственником, тибетским волком, хотя в локусе AMOT таких доказательств не обнаружено (Miao et al., 2017).
Еще одной классической адаптацией к окружающей среде является адаптация африканских собак к малярии, широко распространенной на африканском континенте.Собаки мигрировали в Африку около 14 000 лет назад. Лю и др. (2018) исследовали генетическую основу адаптации к тропической африканской среде посредством анализа всего генома аборигенных африканских собак из Нигерии. Было обнаружено, что среди положительно отобранных генов, связанных с иммунитетом, ангиогенезом, защитой от ультрафиолета, а также секрецией и чувствительностью к инсулину, ген ADGRE1 придает африканским собакам защиту от инфекции Plasmodium (Liu et al. , 2018).Этот ген также связан с иммунным ответом человека на малярию (Kariuki et al., 2013). Таким образом, эти результаты дают новые сведения о конвергентной адаптации и эволюции между людьми и собаками.
, 2018).Этот ген также связан с иммунным ответом человека на малярию (Kariuki et al., 2013). Таким образом, эти результаты дают новые сведения о конвергентной адаптации и эволюции между людьми и собаками.
ФЕНОТИПИЧЕСКОЕ РАЗНООБРАЗИЕ И ОТЕЧЕСТВЕННОЕ НАСЛЕДИЕ СОВРЕМЕННЫХ ПОРОД
С развитием человеческого общества и продвижением промышленной революции потребности людей становились все более разнообразными, а разведение болот было положительно выбрано для двух основных функций. Во-первых, определенных собак разводили для выполнения определенных задач, чтобы помочь своим спутникам-людям, исходя из их особого поведения и функций.Во-вторых, других собак разводили как декоративных питомцев, чтобы интегрировать в человеческие семьи. В настоящее время насчитывается более 400 пород собак, обладающих различными морфологическими и поведенческими характеристиками. Здесь мы рассматриваем их фенотипическое разнообразие, исследуя поведенческие и морфологические фенотипы отдельно.
Поведенческие фенотипы
Породы собак подразделяются на семь групп в соответствии с их охранными, пастушьими, послушными и охотничьими способностями: т. е. охотничьи собаки, рабочие собаки, пастушьи собаки, гончие, терьеры и собаки-компаньоны.Эти поведенческие черты закрепились внутри породы благодаря интенсивному искусственному отбору и считаются наследуемыми, поскольку гибридные собаки демонстрируют промежуточное поведение от своих чистокровных родителей. Обследование более 10 000 особей немецких овчарок и ротвейлеров выявило генетический вклад в 16 оцененных поведенческих черт (Saetre et al., 2006). Дальнейший геномный анализ также показал, что дофаминовый рецептор связан с агрессивным поведением среди собак (Ito et al., 2004). Компульсивное расстройство собак (CCD) распространено у многих пород и демонстрирует механизм, аналогичный механизму человеческого обсессивно-компульсивного расстройства (OCD).Ген кадгерина 2 ( CDh3 ) является решающим фактором, связанным с CCD у доберман-пинчеров (Dodman et al., 2010; Tang et al., 2014). Интересно, что варианты в гене CDh3 также могут способствовать развитию ОКР и синдрома Туретта у людей (Moya et al.
е. охотничьи собаки, рабочие собаки, пастушьи собаки, гончие, терьеры и собаки-компаньоны.Эти поведенческие черты закрепились внутри породы благодаря интенсивному искусственному отбору и считаются наследуемыми, поскольку гибридные собаки демонстрируют промежуточное поведение от своих чистокровных родителей. Обследование более 10 000 особей немецких овчарок и ротвейлеров выявило генетический вклад в 16 оцененных поведенческих черт (Saetre et al., 2006). Дальнейший геномный анализ также показал, что дофаминовый рецептор связан с агрессивным поведением среди собак (Ito et al., 2004). Компульсивное расстройство собак (CCD) распространено у многих пород и демонстрирует механизм, аналогичный механизму человеческого обсессивно-компульсивного расстройства (OCD).Ген кадгерина 2 ( CDh3 ) является решающим фактором, связанным с CCD у доберман-пинчеров (Dodman et al., 2010; Tang et al., 2014). Интересно, что варианты в гене CDh3 также могут способствовать развитию ОКР и синдрома Туретта у людей (Moya et al. , 2013). Более того, популяционно-генетический анализ показал, что CDh3 влияет на кружение у бельгийских малинуа (Cao et al., 2014).
, 2013). Более того, популяционно-генетический анализ показал, что CDh3 влияет на кружение у бельгийских малинуа (Cao et al., 2014).
В дополнение к породным чертам, упомянутым выше, считается, что поведенческая трансформация от страха-агрессивности к приручению была предпосылкой для первоначального одомашнивания собаки.Эксперимент на чернобурых лисицах реконструировал эту поведенческую трансформацию в модели одомашнивания (Lindberg et al., 2005; Saetre et al., 2004), и было обнаружено, что с этим изменением связано несколько генетических локусов (Kukekova et al., 2011; Spady & Острандер, 2007). Однако интенсивное разведение и селекция собак произошли совсем недавно, что привело к чрезвычайно маленькому генофонду, что могло привести к неправильному представлению о генетических изменениях, связанных с трансформацией поведения. Несколько исследований были сосредоточены на китайской аборигенной собаке, беспородной собаке, живущей в первоначальном центре Азии, которая демонстрирует наиболее сильное генетическое разнообразие и, как предполагается, наиболее близка к первичной собаке. Геномный анализ этой беспородной собаки выявил генетическую основу перестройки мозга собаки и конвергентной эволюции между собаками и людьми (Li et al., 2013, 2014a; Lindberg et al., 2005; Saetre et al., 2004; Wang et al. ., 2013).
Геномный анализ этой беспородной собаки выявил генетическую основу перестройки мозга собаки и конвергентной эволюции между собаками и людьми (Li et al., 2013, 2014a; Lindberg et al., 2005; Saetre et al., 2004; Wang et al. ., 2013).
Морфологическое разнообразие
Генетические механизмы, лежащие в основе морфологических различий между собаками, были глубоко изучены, поскольку морфологические признаки легко наблюдать и оценивать. Параллельные исследования морфологических признаков модельных животных, таких как мыши и люди, также выявили генетические основы морфологического разнообразия у собак.
Размер тела, один из наиболее важных количественных признаков, находящихся под пристальным вниманием эволюции, различается у собак больше, чем у любых других наземных млекопитающих. Например, кане-корсо и йоркшир демонстрируют 30-кратную разницу в размерах тела (Shearin & Ostrander, 2010a). Исследования на португальской водяной собаке выявили инсулиноподобный фактор роста 1 ( IGF1 ) в качестве гена, контролирующего размер тела (Sutter et al. , 2007), а также у мышей и людей (Baker et al., 1993; Woods). и другие., 1996, 1997). Варианты фактора роста фибробластов 4 ( FGF4 ) являются причиной коротких ног у многих пород и коррелируют с хондродисплазией (Parker et al., 2009). В отличие от результатов ассоциативного картирования количественных признаков у людей и одомашненных растений, простая генетическая архитектура, по-видимому, лежит в основе морфологической изменчивости собак (Boyko et al., 2010). Например, интегративное исследование нескольких пород показало, что шесть генов ( GHR , HMGA2 , SMAD2 , STC2 , IGF1 и IGF1R ) объясняют почти половину вариаций в породах со стандартным размером тела. вес (BSW) <41 кг (Rimbault et al., 2013). Более того, исследование Х-хромосомы выявило три гена ( IRS4 , IGSF1 и ACSL4 ), отвечающие за изменение размера тела у собак с массой тела >41 кг (Plassais et al., 2017).
, 2007), а также у мышей и людей (Baker et al., 1993; Woods). и другие., 1996, 1997). Варианты фактора роста фибробластов 4 ( FGF4 ) являются причиной коротких ног у многих пород и коррелируют с хондродисплазией (Parker et al., 2009). В отличие от результатов ассоциативного картирования количественных признаков у людей и одомашненных растений, простая генетическая архитектура, по-видимому, лежит в основе морфологической изменчивости собак (Boyko et al., 2010). Например, интегративное исследование нескольких пород показало, что шесть генов ( GHR , HMGA2 , SMAD2 , STC2 , IGF1 и IGF1R ) объясняют почти половину вариаций в породах со стандартным размером тела. вес (BSW) <41 кг (Rimbault et al., 2013). Более того, исследование Х-хромосомы выявило три гена ( IRS4 , IGSF1 и ACSL4 ), отвечающие за изменение размера тела у собак с массой тела >41 кг (Plassais et al., 2017).
Форма черепа — еще одна важная морфологическая черта, которая, как оказалось, очень разнообразна среди пород собак. Анализ формы черепа среди 374 собак выявил SMOC2 как ключевой ген, объясняющий примерно 36% вариаций длины морды (Marchant et al., 2017). Предыдущее исследование ассоциаций морфологии черепа также определило BMP3 как причинный ген, влияющий на развитие черепа и связанный с брахицефалией у некоторых пород, таких как мопс и бульдог (Schoenebeck et al., 2012).
Анализ формы черепа среди 374 собак выявил SMOC2 как ключевой ген, объясняющий примерно 36% вариаций длины морды (Marchant et al., 2017). Предыдущее исследование ассоциаций морфологии черепа также определило BMP3 как причинный ген, влияющий на развитие черепа и связанный с брахицефалией у некоторых пород, таких как мопс и бульдог (Schoenebeck et al., 2012).
Собаки также демонстрируют огромное разнообразие шерсти с тех пор, как они отделились от своего предка серого волка. Полногеномный ассоциативный анализ 80 пород собак выявил три гена ( RSPO2 , FGF5 и KRT71 ), которые влияют на рост, длину и курчавость шерсти (Cadieu et al., 2009). Кроме того, считается, что варианты в семействе транскрипционных факторов forkhead box ( FOX13 ), которые влияют на развитие эктодермы, ответственны за отсутствие шерсти у мексиканских и перуанских бесшерстных собак и китайских хохлатых собак (Drogemuller et al., 2008).
Считается, что три гена, а именно агути ( ASIP ), рецептор меланокортина 1 ( MC1R ) и бета-дефензин 103 ( CBD103 ), кодируют систему лиганд-рецептор, которая контролирует переключение типа пигмента у собак. (Кандилл и др., 2007). Предполагается, что последний ген ( CBD103 ), ранее признанный компонентом врожденного иммунитета у большинства позвоночных, играет роль в функциях, связанных с меланокортиновым путем у домашних собак. Что еще более интересно, североамериканские волки получили аллель CBD103 , что привело к фенотипу черного окраса, в результате генетической интрогрессии от собак более 500 лет назад (Anderson et al., 2009). Кроме того, считается, что гены TYRP1 , MITF и SILV влияют на такие фенотипы, как коричневый цвет, белая пятнистость и мраморный окрас (Clark et al., 2006; Карлссон и др., 2007 г.; Шмутц и др., 2002). Подробную информацию о аллельных паттернах генов, участвующих в окраске шерсти, и связанных с ними фенотипах можно найти в предыдущем обзоре (Schmutz & Berryere, 2007).
(Кандилл и др., 2007). Предполагается, что последний ген ( CBD103 ), ранее признанный компонентом врожденного иммунитета у большинства позвоночных, играет роль в функциях, связанных с меланокортиновым путем у домашних собак. Что еще более интересно, североамериканские волки получили аллель CBD103 , что привело к фенотипу черного окраса, в результате генетической интрогрессии от собак более 500 лет назад (Anderson et al., 2009). Кроме того, считается, что гены TYRP1 , MITF и SILV влияют на такие фенотипы, как коричневый цвет, белая пятнистость и мраморный окрас (Clark et al., 2006; Карлссон и др., 2007 г.; Шмутц и др., 2002). Подробную информацию о аллельных паттернах генов, участвующих в окраске шерсти, и связанных с ними фенотипах можно найти в предыдущем обзоре (Schmutz & Berryere, 2007).
Домашнее наследие
В процессе одомашнивания собак как ослабление селективного давления, так и накопление вредных мутаций из-за эффекта автостопа вызвали домашнее наследие. С одной стороны, селекционеры сосредоточились на очистке сортов и игнорировали устойчивость чистопородных собак к болезням.Сокращение эффективного размера популяции, в основном вызванное очищением породы, увеличило вероятность фиксации многих слабовредных мутаций, расположенных вблизи функциональных локусов, в условиях сильного искусственного отбора (Cruz et al., 2008). С другой стороны, выборочно сохраненные плейотропные гены повышали риск генетических заболеваний. Например, нейросенсорная глухота, связанная с окраской шерсти, широко описана у многих пород собак (Webb & Cullen, 2010), а брахицефалия (плоская морда) обычно наблюдается у таких пород, как английский бульдог, французский бульдог, мопс, бостонский терьер, Пекинес, боксер, ши-тцу и кавалер-кинг-чарльз-спаниель (Dupre & Heidenreich, 2016).Кроме того, у собак часто наблюдаются постоянное кружение, болезнь Альцгеймера и эпилепсия, которые влияют на физическое и психическое благополучие (De Risio et al., 2015; Dewey et al., 2019; Moon-Fanelli & Dodman, 1998).
С одной стороны, селекционеры сосредоточились на очистке сортов и игнорировали устойчивость чистопородных собак к болезням.Сокращение эффективного размера популяции, в основном вызванное очищением породы, увеличило вероятность фиксации многих слабовредных мутаций, расположенных вблизи функциональных локусов, в условиях сильного искусственного отбора (Cruz et al., 2008). С другой стороны, выборочно сохраненные плейотропные гены повышали риск генетических заболеваний. Например, нейросенсорная глухота, связанная с окраской шерсти, широко описана у многих пород собак (Webb & Cullen, 2010), а брахицефалия (плоская морда) обычно наблюдается у таких пород, как английский бульдог, французский бульдог, мопс, бостонский терьер, Пекинес, боксер, ши-тцу и кавалер-кинг-чарльз-спаниель (Dupre & Heidenreich, 2016).Кроме того, у собак часто наблюдаются постоянное кружение, болезнь Альцгеймера и эпилепсия, которые влияют на физическое и психическое благополучие (De Risio et al., 2015; Dewey et al., 2019; Moon-Fanelli & Dodman, 1998). Таким образом, более подробное и надежное понимание геномного состава собак требуется для текущих стратегий разведения, чтобы удовлетворить как меняющиеся потребности человека, так и растущие этические проблемы в отношении благополучия собак.
Таким образом, более подробное и надежное понимание геномного состава собак требуется для текущих стратегий разведения, чтобы удовлетворить как меняющиеся потребности человека, так и растущие этические проблемы в отношении благополучия собак.
ВЫВОДЫ
С развитием технологий секвенирования и снижением связанных с этим затрат, высококачественные данные секвенирования теперь могут обеспечить гораздо более глубокое понимание эволюционной истории собак.Кроме того, улучшение обмена данными в сообществах исследователей собак помогло выяснить генетические механизмы, лежащие в основе наследственности и изменчивости. Например, в рамках Международного проекта Dog10K Genomes Project, который стартовал в 2015 году, были собраны огромные данные по омике пород собак со всего мира. База данных SNP генома собаки (DoGSD) также содержит информацию о различиях между геномами собак и волков (Bai et al., 2015). В дополнение к геномным базам данных международные клубы собаководов также предоставили фенотипическую информацию о более чем 400 породах собак (Ostrander et al. , 2017). Интегрированный ресурс iDog не только содержит всю упомянутую выше информацию (Tang et al., 2019), которая включает в себя инструменты выравнивания последовательностей и браузеры генома, но также может предоставить глобальному сообществу собаководов сборки геномных последовательностей, вариации генома, фенотипы/заболевания. признаки, профили экспрессии генов, онтология генов и информация о гомологах генов. Эти ресурсы будут способствовать нашему пониманию эволюционных и генетических механизмов домашних собак и помогут выяснить генетические основы генетических заболеваний человека.Наконец, как лучший друг человека, собаки заслуживают большего внимания и ответственности за свое благополучие. Владельцы собак должны приобрести профессиональные знания о различных физических состояниях своего компаньона. Ветеринарные ассоциации также должны помочь с введением новых политик, указов, правил и позитивных гуманных решений в интересах собак.
, 2017). Интегрированный ресурс iDog не только содержит всю упомянутую выше информацию (Tang et al., 2019), которая включает в себя инструменты выравнивания последовательностей и браузеры генома, но также может предоставить глобальному сообществу собаководов сборки геномных последовательностей, вариации генома, фенотипы/заболевания. признаки, профили экспрессии генов, онтология генов и информация о гомологах генов. Эти ресурсы будут способствовать нашему пониманию эволюционных и генетических механизмов домашних собак и помогут выяснить генетические основы генетических заболеваний человека.Наконец, как лучший друг человека, собаки заслуживают большего внимания и ответственности за свое благополучие. Владельцы собак должны приобрести профессиональные знания о различных физических состояниях своего компаньона. Ветеринарные ассоциации также должны помочь с введением новых политик, указов, правил и позитивных гуманных решений в интересах собак.
КОНКУРИРУЮЩИЕ ИНТЕРЕСЫ
Авторы заявляют, что у них нет конкурирующих интересов.
АВТОРСКИЕ ВКЛАДЫ
Y.L предложил исследовательские идеи и отредактировал рукопись. З.З. и С.К. просмотрел литературу и составил рукопись. Все авторы прочитали и одобрили окончательный вариант рукописи.
Заявление о финансировании
Эта работа была поддержана Программой Второй научной экспедиции и исследований Тибетского нагорья (STEP) (2019QZKK0501), Программой стратегических приоритетных исследований Китайской академии наук (XDA2004010302) и Национальным фондом естественных наук Китая (31860305). ).Ю.Л. был поддержан Фондом молодых академических и технических лидеров провинции Юньнань
Ссылки
1. Андерсон Т.М., фон Холдт Б.М., Кандиль С.И., Мусиани М., Греко К., Шталер Д.Р., Смит Д.В., Падхукасахасрам Б., Рэнди Э., Леонард Дж.А., Бустаманте К.Д., Острандер Э.А., Тан Х., Уэйн Р.К., Барш Г.С. Молекулярная и эволюционная история меланизма у серых волков Северной Америки. Наука. 2009;323(5919):1339–1343. doi: 10.1126/science.1165448. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]2. Арендт М., Кэрнс К.М., Баллард Дж.В.О., Саволайнен П., Аксельссон Э. Адаптация диеты у собак отражает распространение доисторического земледелия. Наследственность (Эдинб) 2016;117(5):301–306. doi: 10.1038/hdy.2016.48. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]3. Арендт М., Фолл Т., Линдблад-Тох К., Аксельссон Э. Активность амилазы связана с количеством копий AMY2B у собак: последствия для одомашнивания собак, диеты и диабета. Генетика животных. 2014;45(5):716–722. doi: 10.1111/возраст.12179. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]4.Аксельссон Э., Ратнакумар А., Арендт М.Л., Макбул К., Вебстер М.Т., Перлоски М., Либерг О., Арнемо Дж.М., Хедхаммар О., Линдблад-Тох К. Геномная характеристика одомашнивания собак показывает адаптацию к диете, богатой крахмалом. Природа. 2013;495(7441):360–364. doi: 10.1038/nature11837. [PubMed] [CrossRef] [Google Scholar]5. Bai B, Zhao WM, Tang BX, Wang YQ, Wang L, Zhang Z, Yang HC, Liu YH, Zhu JW, Irwin DM, Wang GD, Zhang YP DoGSD: база данных SNP генома собаки и волка.
Арендт М., Кэрнс К.М., Баллард Дж.В.О., Саволайнен П., Аксельссон Э. Адаптация диеты у собак отражает распространение доисторического земледелия. Наследственность (Эдинб) 2016;117(5):301–306. doi: 10.1038/hdy.2016.48. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]3. Арендт М., Фолл Т., Линдблад-Тох К., Аксельссон Э. Активность амилазы связана с количеством копий AMY2B у собак: последствия для одомашнивания собак, диеты и диабета. Генетика животных. 2014;45(5):716–722. doi: 10.1111/возраст.12179. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]4.Аксельссон Э., Ратнакумар А., Арендт М.Л., Макбул К., Вебстер М.Т., Перлоски М., Либерг О., Арнемо Дж.М., Хедхаммар О., Линдблад-Тох К. Геномная характеристика одомашнивания собак показывает адаптацию к диете, богатой крахмалом. Природа. 2013;495(7441):360–364. doi: 10.1038/nature11837. [PubMed] [CrossRef] [Google Scholar]5. Bai B, Zhao WM, Tang BX, Wang YQ, Wang L, Zhang Z, Yang HC, Liu YH, Zhu JW, Irwin DM, Wang GD, Zhang YP DoGSD: база данных SNP генома собаки и волка. Исследование нуклеиновых кислот. 2015;43(D1):D777–D783. дои: 10.1093/нар/гку1174. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]6. Бейкер Дж., Лю Дж. П., Робертсон Э. Дж., Эфстратиадис Роль инсулиноподобных факторов роста в эмбриональном и постнатальном росте. Клетка. 1993;75(1):73–82. doi: 10.1016/S0092-8674(05)80085-6. [PubMed] [CrossRef] [Google Scholar]7. Бойко А.Р., Бойко Р.Х., Бойко К.М., Паркер Х.Г., Кастельхано М., Кори Л., Дегенхардт Дж.Д., Аутон А., Хедимби М., Китио Р., Острандер Э.А., Шонебек Дж., Тодхантер Р.Дж., Джонс П., Бустаманте К.Д. Сложная структура населения в африканской деревне собаки и его последствия для выведения истории одомашнивания собак.Труды Национальной академии наук Соединенных Штатов Америки. 2009;106(33):13903–13908. doi: 10.1073/pnas.09106. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]8. Бойко А.Р., Куиньон П., Ли Л., Шёнебек Дж.Дж., Дегенхардт Дж.Д., Ломюллер К.Е., Чжао К., Брисбин А., Паркер Х.Г., фон Холдт Б.М., Каргилл М.
Исследование нуклеиновых кислот. 2015;43(D1):D777–D783. дои: 10.1093/нар/гку1174. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]6. Бейкер Дж., Лю Дж. П., Робертсон Э. Дж., Эфстратиадис Роль инсулиноподобных факторов роста в эмбриональном и постнатальном росте. Клетка. 1993;75(1):73–82. doi: 10.1016/S0092-8674(05)80085-6. [PubMed] [CrossRef] [Google Scholar]7. Бойко А.Р., Бойко Р.Х., Бойко К.М., Паркер Х.Г., Кастельхано М., Кори Л., Дегенхардт Дж.Д., Аутон А., Хедимби М., Китио Р., Острандер Э.А., Шонебек Дж., Тодхантер Р.Дж., Джонс П., Бустаманте К.Д. Сложная структура населения в африканской деревне собаки и его последствия для выведения истории одомашнивания собак.Труды Национальной академии наук Соединенных Штатов Америки. 2009;106(33):13903–13908. doi: 10.1073/pnas.09106. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]8. Бойко А.Р., Куиньон П., Ли Л., Шёнебек Дж.Дж., Дегенхардт Дж.Д., Ломюллер К.Е., Чжао К., Брисбин А., Паркер Х.Г., фон Холдт Б.М., Каргилл М. , Аутон А., Рейнольдс А., Элькалун А.Г., Кастельхано М., Мошер Д.С., Саттер Н.Б. , Johnson GS, Novembre J, Hubisz MJ, Siepel A, Wayne RK, Bustamante CD, Ostrander EA В основе морфологической изменчивости собак лежит простая генетическая архитектура.Биология PLoS. 2010;8(8):e1000451. doi: 10.1371/journal.pbio.1000451. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]9. Кадье Э., Нефф М.В., Куиньон П., Уолш К., Чейз К., Паркер Х.Г., ФонХолдт Б.М., Рю А., Бойко А., Байерс А., Вонг А., Мошер Д.С., Элкалун А.Г., Спади Т.К., АндреК, Ларк К.Г., Каргилл М., Бустаманте К.Д., Уэйн Р.К., Острандер Э.А. Изменчивость шерсти домашней собаки определяется вариантами трех генов. Наука. 2009;326(5949):150–153. doi: 10.1126/science.1177808. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]10.Candille SI, Kaelin CB, Cattanach BM, Yu B, Thompson DA, Nix MA, Kerns JA, Schmutz SM, Millhauser GL, Barsh GS Мутация β-дефенсина вызывает черный цвет шерсти у домашних собак. Наука. 2007;318(5855):1418–1423.
, Аутон А., Рейнольдс А., Элькалун А.Г., Кастельхано М., Мошер Д.С., Саттер Н.Б. , Johnson GS, Novembre J, Hubisz MJ, Siepel A, Wayne RK, Bustamante CD, Ostrander EA В основе морфологической изменчивости собак лежит простая генетическая архитектура.Биология PLoS. 2010;8(8):e1000451. doi: 10.1371/journal.pbio.1000451. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]9. Кадье Э., Нефф М.В., Куиньон П., Уолш К., Чейз К., Паркер Х.Г., ФонХолдт Б.М., Рю А., Бойко А., Байерс А., Вонг А., Мошер Д.С., Элкалун А.Г., Спади Т.К., АндреК, Ларк К.Г., Каргилл М., Бустаманте К.Д., Уэйн Р.К., Острандер Э.А. Изменчивость шерсти домашней собаки определяется вариантами трех генов. Наука. 2009;326(5949):150–153. doi: 10.1126/science.1177808. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]10.Candille SI, Kaelin CB, Cattanach BM, Yu B, Thompson DA, Nix MA, Kerns JA, Schmutz SM, Millhauser GL, Barsh GS Мутация β-дефенсина вызывает черный цвет шерсти у домашних собак. Наука. 2007;318(5855):1418–1423. doi: 10.1126/science.1147880. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]11. Cao X, Irwin DM, Liu YH, Cheng LG, Wang L, Wang GD, Zhang YP Балансирующий отбор по CDh3 может быть связан с поведенческими особенностями бельгийского малинуа. ПЛОС Один. 2014;9(10):e110075. doi: 10.1371/журнал.поне.0110075. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]12. Clark LA, Wahl JM, Rees CA, Murphy KE Вставка ретротранспозона в SILV ответственна за паттерн мраморного окраса у домашней собаки. Труды Национальной академии наук Соединенных Штатов Америки. 2006;103(5):1376–1381. doi: 10.1073/pnas.0506940103. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]13. Круз Ф., Вила С., Вебстер М.Т. Наследие одомашнивания: накопление вредных мутаций в геноме собаки.Молекулярная биология и эволюция. 2008;25(11):2331–2336. doi: 10.1093/molbev/msn177. [PubMed] [CrossRef] [Google Scholar] 14. Де Рисио Л., Бхатти С., Муньяна К., Пендерис Дж., Штейн В., Типольд А., Берендт М., Фаркуар Р.
doi: 10.1126/science.1147880. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]11. Cao X, Irwin DM, Liu YH, Cheng LG, Wang L, Wang GD, Zhang YP Балансирующий отбор по CDh3 может быть связан с поведенческими особенностями бельгийского малинуа. ПЛОС Один. 2014;9(10):e110075. doi: 10.1371/журнал.поне.0110075. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]12. Clark LA, Wahl JM, Rees CA, Murphy KE Вставка ретротранспозона в SILV ответственна за паттерн мраморного окраса у домашней собаки. Труды Национальной академии наук Соединенных Штатов Америки. 2006;103(5):1376–1381. doi: 10.1073/pnas.0506940103. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]13. Круз Ф., Вила С., Вебстер М.Т. Наследие одомашнивания: накопление вредных мутаций в геноме собаки.Молекулярная биология и эволюция. 2008;25(11):2331–2336. doi: 10.1093/molbev/msn177. [PubMed] [CrossRef] [Google Scholar] 14. Де Рисио Л., Бхатти С., Муньяна К., Пендерис Дж., Штейн В., Типольд А., Берендт М., Фаркуар Р. , Фишер А., Лонг С., Мандигерс П.Дж., Матиасек К., Пакер Р.М., Пакозди А., Паттерсон Н., Платт С., Поделл M, Potschka H, Batlle MP, Rusbridge C, Volk HA Консенсусное предложение Международной ветеринарной рабочей группы по эпилепсии: диагностический подход к эпилепсии у собак. Ветеринарные исследования BMC. 2015;11:148. дои: 10.1186/с12917-015-0462-1. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]15. Dewey CW, Davies ES, Xie H, Wakshlag JJ Когнитивная дисфункция собак: патофизиология, диагностика и лечение. Ветеринарные клиники Северной Америки-Практика мелких животных. 2019;49(3):477–499. doi: 10.1016/j.cvsm.2019.01.013. [PubMed] [CrossRef] [Google Scholar] 16. Дин З.Л., Оскарссон М., Ардалан А., Энглби Х., Дальгрен Л.Г., Тепели С., Киркнесс Э., Саволайнен П., Чжан Ю.П. Происхождение домашних собак в Юго-Восточной Азии подтверждается анализом ДНК Y-хромосомы.Наследственность. 2012;108(5):507–514. doi: 10.1038/hdy.2011.114. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]17.
, Фишер А., Лонг С., Мандигерс П.Дж., Матиасек К., Пакер Р.М., Пакозди А., Паттерсон Н., Платт С., Поделл M, Potschka H, Batlle MP, Rusbridge C, Volk HA Консенсусное предложение Международной ветеринарной рабочей группы по эпилепсии: диагностический подход к эпилепсии у собак. Ветеринарные исследования BMC. 2015;11:148. дои: 10.1186/с12917-015-0462-1. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]15. Dewey CW, Davies ES, Xie H, Wakshlag JJ Когнитивная дисфункция собак: патофизиология, диагностика и лечение. Ветеринарные клиники Северной Америки-Практика мелких животных. 2019;49(3):477–499. doi: 10.1016/j.cvsm.2019.01.013. [PubMed] [CrossRef] [Google Scholar] 16. Дин З.Л., Оскарссон М., Ардалан А., Энглби Х., Дальгрен Л.Г., Тепели С., Киркнесс Э., Саволайнен П., Чжан Ю.П. Происхождение домашних собак в Юго-Восточной Азии подтверждается анализом ДНК Y-хромосомы.Наследственность. 2012;108(5):507–514. doi: 10.1038/hdy.2011.114. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]17. Добни К., Ларсон Г. Генетика и одомашнивание животных: новое окно в неуловимый процесс. Журнал зоологии. 2006;269(2):261–271. [Google Академия] 18. Dodman NH, Karlsson EK, Moon-Fanelli A, Galdzicka M, Perloski M, Shuster L, Lindblad-Toh K, Ginns EI Локус собачьей хромосомы 7 придает предрасположенность к компульсивному расстройству. Молекулярная психиатрия. 2010;15(1):8–10. doi: 10.1038/mp.2009.111. [PubMed] [CrossRef] [Google Scholar]
Добни К., Ларсон Г. Генетика и одомашнивание животных: новое окно в неуловимый процесс. Журнал зоологии. 2006;269(2):261–271. [Google Академия] 18. Dodman NH, Karlsson EK, Moon-Fanelli A, Galdzicka M, Perloski M, Shuster L, Lindblad-Toh K, Ginns EI Локус собачьей хромосомы 7 придает предрасположенность к компульсивному расстройству. Молекулярная психиатрия. 2010;15(1):8–10. doi: 10.1038/mp.2009.111. [PubMed] [CrossRef] [Google Scholar]19. Doebley J. 1989. Изоферментные доказательства и эволюция сельскохозяйственных растений. В : Солтис Д.Е., Солтис П.С., Дадли Т.Р. (ред.). Изоферменты в биологии растений. Дордрехт: Спрингер.
20. Drogemuller C, Karlsson EK, Hytönen MK, Perloski M, Dolf G, Sainio K, Lohi H, Lindblad-Toh K, Leeb T Мутация у бесшерстных собак связана с FOXI3 в развитии эктодермы. Наука. 2008;321(5895):1462–1462. doi: 10.1126/science.1162525. [PubMed] [CrossRef] [Google Scholar] 21.Dupre G, Heidenreich D Брахицефальный синдром. Ветеринарные клиники Северной Америки-Практика мелких животных. 2016;46(4):691–707. doi: 10.1016/j.cvsm.2016.02.002. [PubMed] [CrossRef] [Google Scholar] 22. Эйр-Уокер А., Гаут Р.Л., Хилтон Х., Фельдман Д.Л., Гаут Б.С. Исследование узкого места, ведущего к одомашниванию кукурузы. Труды Национальной академии наук Соединенных Штатов Америки. 1998;95(8):4441–4446. doi: 10.1073/pnas.95.8.4441. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]23.Fan Z, Silva P, Gronau I, Wang S, Armero AS, Schweizer RM, Ramirez O, Pollinger J, Galaverni M, Ortega Del-Vecchyo D, Du L, Zhang W, Zhang Z, Xing J, Vila C, Marques- Бонет Т., Годиньо Р., Юэ Б., Уэйн Р.К. Общемировые закономерности геномной изменчивости и примеси у серых волков. Геномные исследования. 2016;26(2):163–73. doi: 10.1101/gr.197517.115. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]24. Франц Л.А., Маллин В.Е., Пионье-Капитан М., Лебрассер О., Оливье М., Перри А., Линдерхольм А., Маттиангели В., Тисдейл М.Д., Димопулос Э.А., Трессе А., Дюффрас М., Маккормик Ф., Бартосевич Л., Гал Э.
2016;46(4):691–707. doi: 10.1016/j.cvsm.2016.02.002. [PubMed] [CrossRef] [Google Scholar] 22. Эйр-Уокер А., Гаут Р.Л., Хилтон Х., Фельдман Д.Л., Гаут Б.С. Исследование узкого места, ведущего к одомашниванию кукурузы. Труды Национальной академии наук Соединенных Штатов Америки. 1998;95(8):4441–4446. doi: 10.1073/pnas.95.8.4441. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]23.Fan Z, Silva P, Gronau I, Wang S, Armero AS, Schweizer RM, Ramirez O, Pollinger J, Galaverni M, Ortega Del-Vecchyo D, Du L, Zhang W, Zhang Z, Xing J, Vila C, Marques- Бонет Т., Годиньо Р., Юэ Б., Уэйн Р.К. Общемировые закономерности геномной изменчивости и примеси у серых волков. Геномные исследования. 2016;26(2):163–73. doi: 10.1101/gr.197517.115. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]24. Франц Л.А., Маллин В.Е., Пионье-Капитан М., Лебрассер О., Оливье М., Перри А., Линдерхольм А., Маттиангели В., Тисдейл М.Д., Димопулос Э.А., Трессе А., Дюффрас М., Маккормик Ф., Бартосевич Л., Гал Э. , Ньергес Э.А., Саблин М.В., Брехард С., Машкур М., Бэлэшеску А., Джиллет Б., Хьюз С., Чессейн О., Хитте С., Винье Д.Д., Добни К., Хенни С., Брэдли Д.Г., Ларсон Г. Геномные и археологические данные свидетельствуют о двойном происхождении домашних собак.Наука. 2016;352(6290):1228–1231. doi: 10.1126/science.aaf3161. [PubMed] [CrossRef] [Google Scholar] 25. Фридман А.Х., Гронау И., Швейцер Р.М., Ортега-дель Веккио Д., Хан Э., Сильва П.М., Галаверни М., Фан З., Маркс П., Лоренте-Галдос Б., Бил Х., Рамирес О., Хормоздиари Ф., Алкан К., Вила К., Сквайр К., Геффен Э., Кусак Дж., Бойко А.Р., Паркер Х.Г., Ли С., Тадиготла В., Уилтон А., Зипель А., Бустаманте К.Д., Харкинс Т.Т., Нельсон С.Ф., Острандер Э.А., Маркес-Бонет Т., Уэйн Р.К., Novembre J Genome секвенирование подчеркивает динамическую раннюю историю собак.Генетика PLoS. 2014;10(1):e1004016. doi: 10.1371/journal.pgen.1004016. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]26. Фридман А.Х., Уэйн Р.К. Расшифровка происхождения собак: от окаменелостей к геномам.
, Ньергес Э.А., Саблин М.В., Брехард С., Машкур М., Бэлэшеску А., Джиллет Б., Хьюз С., Чессейн О., Хитте С., Винье Д.Д., Добни К., Хенни С., Брэдли Д.Г., Ларсон Г. Геномные и археологические данные свидетельствуют о двойном происхождении домашних собак.Наука. 2016;352(6290):1228–1231. doi: 10.1126/science.aaf3161. [PubMed] [CrossRef] [Google Scholar] 25. Фридман А.Х., Гронау И., Швейцер Р.М., Ортега-дель Веккио Д., Хан Э., Сильва П.М., Галаверни М., Фан З., Маркс П., Лоренте-Галдос Б., Бил Х., Рамирес О., Хормоздиари Ф., Алкан К., Вила К., Сквайр К., Геффен Э., Кусак Дж., Бойко А.Р., Паркер Х.Г., Ли С., Тадиготла В., Уилтон А., Зипель А., Бустаманте К.Д., Харкинс Т.Т., Нельсон С.Ф., Острандер Э.А., Маркес-Бонет Т., Уэйн Р.К., Novembre J Genome секвенирование подчеркивает динамическую раннюю историю собак.Генетика PLoS. 2014;10(1):e1004016. doi: 10.1371/journal.pgen.1004016. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]26. Фридман А.Х., Уэйн Р.К. Расшифровка происхождения собак: от окаменелостей к геномам. Ежегодный обзор биологических наук о животных. 2017;5:281–307. doi: 10.1146/annurev-animal-022114-110937. [PubMed] [CrossRef] [Google Scholar] 27. Гоу С, Ван З, Ли Н, Цю Ф, Сю З, Ян Д.В., Ян С.Л., Цзя Дж., Конг С.Ю., Вэй Ч.Х., Лу С.С., Лянь Л.С., Ву С.Х., Ван С.Ю., Ли Г.З., Ма Т., Цзян К. , Чжао X, Ян Дж. К., Лю Б. Х., Вэй Д. К., Ли Х, Ян Дж. Ф., Ян Ю. Л., Чжао Г. Ю., Дун ХХ, Ли М. Л., Дэн В. Д., Ленг Дж., Вэй К. С., Ван С., Мао Х. М., Чжан Х., Дин GH, Li YX Полногеномное секвенирование шести пород собак, живших на постоянной высоте, выявило адаптацию к высокогорной гипоксии.Геномные исследования. 2014;24(8):1308–1315. doi: 10.1101/gr.171876.113. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]28. Ito H, Nara H, Inoue-Murayama M, Shimada MK, Koshimura A, Ueda Y, Kitagawa H, Takeuchi Y, Mori Y, Murayama Y, Morita M, Iwasaki T, Ôta K, Tanabe Y, Ito S Частотное распределение аллелей экзон III и I гена дофаминового рецептора D4 собак у 23 пород. Журнал ветеринарной медицины. 2004;66(7):815–820.
Ежегодный обзор биологических наук о животных. 2017;5:281–307. doi: 10.1146/annurev-animal-022114-110937. [PubMed] [CrossRef] [Google Scholar] 27. Гоу С, Ван З, Ли Н, Цю Ф, Сю З, Ян Д.В., Ян С.Л., Цзя Дж., Конг С.Ю., Вэй Ч.Х., Лу С.С., Лянь Л.С., Ву С.Х., Ван С.Ю., Ли Г.З., Ма Т., Цзян К. , Чжао X, Ян Дж. К., Лю Б. Х., Вэй Д. К., Ли Х, Ян Дж. Ф., Ян Ю. Л., Чжао Г. Ю., Дун ХХ, Ли М. Л., Дэн В. Д., Ленг Дж., Вэй К. С., Ван С., Мао Х. М., Чжан Х., Дин GH, Li YX Полногеномное секвенирование шести пород собак, живших на постоянной высоте, выявило адаптацию к высокогорной гипоксии.Геномные исследования. 2014;24(8):1308–1315. doi: 10.1101/gr.171876.113. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]28. Ito H, Nara H, Inoue-Murayama M, Shimada MK, Koshimura A, Ueda Y, Kitagawa H, Takeuchi Y, Mori Y, Murayama Y, Morita M, Iwasaki T, Ôta K, Tanabe Y, Ito S Частотное распределение аллелей экзон III и I гена дофаминового рецептора D4 собак у 23 пород. Журнал ветеринарной медицины. 2004;66(7):815–820. doi: 10.1292/jvms.66.815. [PubMed] [CrossRef] [Google Scholar] 29.Kariuki SM, Rockett K, Clark TG, Reyburn H, Agbenyega T, Taylor TE, Birbeck GL, Williams TN, Newton CRJC Генетический риск острых судорог у африканских детей с малярией falciparum. Эпилепсия. 2013;54(6):990–1001. doi: 10.1111/эпи.12173. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]30. Карлссон Э.К., Барановска И., Уэйд С.М., Салмон Хиллбертц Н.Х., Зоди М.С., Андерсон Н., Биаги Т.М., Паттерсон Н., Пилберг Г.Р., Кульбокас Э.Дж., Комсток К.Е., Келлер Э.Т., Месиров Д.П., фон Эйлер Х., Кампе О., Хедхаммар А., Ландер Э.С., Андерссон Г., Андерссон Л., Линдблад-Тох К. Эффективное картирование менделевских признаков у собак посредством полногеномной ассоциации.Генетика природы. 2007;39(11):1321–1328. doi: 10.1038/ng.2007.10. [PubMed] [CrossRef] [Google Scholar] 31. Кукекова А.В., Трут Л.Н., Чейз К., Харламова А.В., Джонсон Дж.Л., Темных С.В., Оськина И.Н., Гулевич Р.Г., Владимирова А.В., Клебанов С., Шепелева Д.В., Шихевич С.
doi: 10.1292/jvms.66.815. [PubMed] [CrossRef] [Google Scholar] 29.Kariuki SM, Rockett K, Clark TG, Reyburn H, Agbenyega T, Taylor TE, Birbeck GL, Williams TN, Newton CRJC Генетический риск острых судорог у африканских детей с малярией falciparum. Эпилепсия. 2013;54(6):990–1001. doi: 10.1111/эпи.12173. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]30. Карлссон Э.К., Барановска И., Уэйд С.М., Салмон Хиллбертц Н.Х., Зоди М.С., Андерсон Н., Биаги Т.М., Паттерсон Н., Пилберг Г.Р., Кульбокас Э.Дж., Комсток К.Е., Келлер Э.Т., Месиров Д.П., фон Эйлер Х., Кампе О., Хедхаммар А., Ландер Э.С., Андерссон Г., Андерссон Л., Линдблад-Тох К. Эффективное картирование менделевских признаков у собак посредством полногеномной ассоциации.Генетика природы. 2007;39(11):1321–1328. doi: 10.1038/ng.2007.10. [PubMed] [CrossRef] [Google Scholar] 31. Кукекова А.В., Трут Л.Н., Чейз К., Харламова А.В., Джонсон Дж.Л., Темных С.В., Оськина И.Н., Гулевич Р.Г., Владимирова А.В., Клебанов С., Шепелева Д.В., Шихевич С. Г., Акланд Г.М., Ларк К.Г. Картирование локусов доместикации лисицы: деконструкция/реконструкция поведенческого фенотипа. Генетика поведения. 2011;41(4):593–606. doi: 10.1007/s10519-010-9418-1. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]32. Леонард Дж. А., Уэйн Р. К., Уилер Дж., Валадес Р., Гильен С., Вила С Доказательства древней ДНК происхождения собак Нового Света из Старого Света.Наука. 2002;298(5598):1613–1616. doi: 10.1126/science.1076980. [PubMed] [CrossRef] [Google Scholar] 33. Ли Ю., фон Холдт Б.М., Рейнольдс А., Бойко А.Р., Уэйн Р.К., Ву Д.Д., Чжан Ю.П. Искусственный отбор генов, экспрессируемых в мозге, во время одомашнивания собак. Молекулярная биология и эволюция. 2013;30(8):1867–1876. doi: 10.1093/molbev/mst088. [PubMed] [CrossRef] [Google Scholar] 34. Ли И, Ван Г.Д., Ван М.С., Ирвин Д.М., Ву Д.Д., Чжан Ю.П. Одомашниванию собаки от волка способствовала повышенная возбуждающая синаптическая пластичность: гипотеза.Геномная биология и эволюция. 2014а;6(11):3115–3121.
Г., Акланд Г.М., Ларк К.Г. Картирование локусов доместикации лисицы: деконструкция/реконструкция поведенческого фенотипа. Генетика поведения. 2011;41(4):593–606. doi: 10.1007/s10519-010-9418-1. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]32. Леонард Дж. А., Уэйн Р. К., Уилер Дж., Валадес Р., Гильен С., Вила С Доказательства древней ДНК происхождения собак Нового Света из Старого Света.Наука. 2002;298(5598):1613–1616. doi: 10.1126/science.1076980. [PubMed] [CrossRef] [Google Scholar] 33. Ли Ю., фон Холдт Б.М., Рейнольдс А., Бойко А.Р., Уэйн Р.К., Ву Д.Д., Чжан Ю.П. Искусственный отбор генов, экспрессируемых в мозге, во время одомашнивания собак. Молекулярная биология и эволюция. 2013;30(8):1867–1876. doi: 10.1093/molbev/mst088. [PubMed] [CrossRef] [Google Scholar] 34. Ли И, Ван Г.Д., Ван М.С., Ирвин Д.М., Ву Д.Д., Чжан Ю.П. Одомашниванию собаки от волка способствовала повышенная возбуждающая синаптическая пластичность: гипотеза.Геномная биология и эволюция. 2014а;6(11):3115–3121. doi: 10.1093/gbe/evu245. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]35. Li Y, Wu DD, Boyko AR, Wang GD, Wu SF, Irwin DM, Zhang YP Изменение популяции выявило высотную адаптацию тибетских мастифов. Молекулярная биология и эволюция. 2014b;31(5):1200–1205. doi: 10.1093/molbev/msu070. [PubMed] [CrossRef] [Google Scholar] 36. Lindberg J, Björnerfeldt S, Saetre P, Svartberg K, Seehuus B, Bakken M, Vilà C, Jazin E Отбор на приручение изменил экспрессию генов мозга у чернобурых лисиц.Текущая биология. 2005;15(22):R915–R916. doi: 10.1016/j.cub.2005.11.009. [PubMed] [CrossRef] [Google Scholar] 37. Лю Ю.Х., Ван Л., Сюй Т., Го Х.М., Ли И., Инь Т.Т., Ян Х.К., Ху И., Адеола А.С., Санке О.Дж., Отецко Н.О., Ван М., Ма Ю.П., Чарльз О.С., Синдинг МХС, Гопалакришнан С., Саманьего Дж.А. , Hansen AJ, Fernandes C, Gaubert P, Budd J, Dawuda PM, Rueness EK, Jiang LB, Zhai WW, Gilbert MTP, Peng MS, Qi XP, Wang GD, Zhang YP Полногеномное секвенирование африканских собак дает представление об адаптации против тропических паразитов.
doi: 10.1093/gbe/evu245. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]35. Li Y, Wu DD, Boyko AR, Wang GD, Wu SF, Irwin DM, Zhang YP Изменение популяции выявило высотную адаптацию тибетских мастифов. Молекулярная биология и эволюция. 2014b;31(5):1200–1205. doi: 10.1093/molbev/msu070. [PubMed] [CrossRef] [Google Scholar] 36. Lindberg J, Björnerfeldt S, Saetre P, Svartberg K, Seehuus B, Bakken M, Vilà C, Jazin E Отбор на приручение изменил экспрессию генов мозга у чернобурых лисиц.Текущая биология. 2005;15(22):R915–R916. doi: 10.1016/j.cub.2005.11.009. [PubMed] [CrossRef] [Google Scholar] 37. Лю Ю.Х., Ван Л., Сюй Т., Го Х.М., Ли И., Инь Т.Т., Ян Х.К., Ху И., Адеола А.С., Санке О.Дж., Отецко Н.О., Ван М., Ма Ю.П., Чарльз О.С., Синдинг МХС, Гопалакришнан С., Саманьего Дж.А. , Hansen AJ, Fernandes C, Gaubert P, Budd J, Dawuda PM, Rueness EK, Jiang LB, Zhai WW, Gilbert MTP, Peng MS, Qi XP, Wang GD, Zhang YP Полногеномное секвенирование африканских собак дает представление об адаптации против тропических паразитов. Молекулярная биология и эволюция. 2018;35(2):287–298. doi: 10.1093/molbev/msx258. [PubMed] [CrossRef] [Google Scholar] 38. Маршан Т.В., Джонсон Э.Дж., Мактейр Л., Джонсон К.И., Гоу А., Люти Т., Куэн Д., Свенсон К., Бермингем М.Л., Дрогемюллер М., Нуссбаумер М., Дэйви М.Г., Аргайл Д.Дж., Пауэлл Р.М., Гильерме С., Ланг Дж., Тер Хаар G, Leeb T, Schwarz T, Mellanby RJ, Clements DN, Schoenebeck JJ Собачья брахицефалия связана с ретротранспозон-опосредованным неправильным сплайсингом SMOC2. Текущая биология. 2017;27(11):1573–1584.doi: 10.1016/j.cub.2017.04.057. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]39. Miao B, Wang Z, Li Y Геномный анализ выявил адаптацию к гипоксии у тибетского мастифа путем интрогрессии серого волка с Тибетского нагорья. Молекулярная биология и эволюция. 2017;34(3):734–743. [PubMed] [Google Scholar]40. Мун-Фанелли А.А., Додман Н.Х. Описание и развитие навязчивой погони за хвостом у терьеров и ответ на лечение кломипрамином. Журнал Американской ветеринарной медицинской ассоциации.
Молекулярная биология и эволюция. 2018;35(2):287–298. doi: 10.1093/molbev/msx258. [PubMed] [CrossRef] [Google Scholar] 38. Маршан Т.В., Джонсон Э.Дж., Мактейр Л., Джонсон К.И., Гоу А., Люти Т., Куэн Д., Свенсон К., Бермингем М.Л., Дрогемюллер М., Нуссбаумер М., Дэйви М.Г., Аргайл Д.Дж., Пауэлл Р.М., Гильерме С., Ланг Дж., Тер Хаар G, Leeb T, Schwarz T, Mellanby RJ, Clements DN, Schoenebeck JJ Собачья брахицефалия связана с ретротранспозон-опосредованным неправильным сплайсингом SMOC2. Текущая биология. 2017;27(11):1573–1584.doi: 10.1016/j.cub.2017.04.057. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]39. Miao B, Wang Z, Li Y Геномный анализ выявил адаптацию к гипоксии у тибетского мастифа путем интрогрессии серого волка с Тибетского нагорья. Молекулярная биология и эволюция. 2017;34(3):734–743. [PubMed] [Google Scholar]40. Мун-Фанелли А.А., Додман Н.Х. Описание и развитие навязчивой погони за хвостом у терьеров и ответ на лечение кломипрамином. Журнал Американской ветеринарной медицинской ассоциации. 1998;212(8):1252–1257. [PubMed] [Google Scholar]41. Moya PR, Dodman NH, Timpano KR, Rubenstein LM, Rana Z, Fried RL, Reichardt LF, Heiman GA, Tischfield JA, King RA, Galdzicka M, Ginns EI, Wendland JR Редкие варианты гена миссенс-нейронального кадгерина (CDh3) при специфических обсессивных состояниях фенотипы компульсивного расстройства и синдрома Туретта. Европейский журнал генетики человека. 2013;21(8):850–854. doi: 10.1038/ejhg.2012.245. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]42. Ни Литлобхейр М., Перри А.Р., Ирвинг-Пиз Э.К., Витт К.Е., Линдерхольм А., Хайле Дж., Лебрассер О., Амин С., Блик Дж., Бойко А.Р., Брейс С., Кортес Ю.Н., Крокфорд С.Дж., Дево А., Димопулос Э.А., Элдридж М. , Энк Дж., Гопалакришнан С., Гори К., Граймс В., Гири Э., Хансен А.Дж., Халм-Биман А., Джонсон Дж., Китчен А., Каспаров А.К., Квон Ю.М., Никольский П.А., Лопе С.П., Манин А., Мартин Т., Мейер М. , Майерс К.Н., Омура М., Руйяр Дж.М., Павлова Е.Ю., Скиулли П., Синдинг М.С., Стракова А., Иванова В.
1998;212(8):1252–1257. [PubMed] [Google Scholar]41. Moya PR, Dodman NH, Timpano KR, Rubenstein LM, Rana Z, Fried RL, Reichardt LF, Heiman GA, Tischfield JA, King RA, Galdzicka M, Ginns EI, Wendland JR Редкие варианты гена миссенс-нейронального кадгерина (CDh3) при специфических обсессивных состояниях фенотипы компульсивного расстройства и синдрома Туретта. Европейский журнал генетики человека. 2013;21(8):850–854. doi: 10.1038/ejhg.2012.245. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]42. Ни Литлобхейр М., Перри А.Р., Ирвинг-Пиз Э.К., Витт К.Е., Линдерхольм А., Хайле Дж., Лебрассер О., Амин С., Блик Дж., Бойко А.Р., Брейс С., Кортес Ю.Н., Крокфорд С.Дж., Дево А., Димопулос Э.А., Элдридж М. , Энк Дж., Гопалакришнан С., Гори К., Граймс В., Гири Э., Хансен А.Дж., Халм-Биман А., Джонсон Дж., Китчен А., Каспаров А.К., Квон Ю.М., Никольский П.А., Лопе С.П., Манин А., Мартин Т., Мейер М. , Майерс К.Н., Омура М., Руйяр Дж.М., Павлова Е.Ю., Скиулли П., Синдинг М.С., Стракова А., Иванова В. В., Видга С., Виллерслев Э., Питулько В.В., Барнс И., Гилберт МТП, Добни К.М., Малхи Р.С., Мерчисон Э.П., Ларсон G, Frantz LAF Эволюционная история собак в Америке.Наука. 2018;361(6397):81–85. doi: 10.1126/science.aao4776. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]43. Оливье М., Трессе А., Франц Лаф, Брехард С., Бэлашеску А., Машкур М., Боронеант А., Пионье-Капитан М., Лебрассер О., Арбогаст Р.М., Бартосевич Л., Дебю К., Рабинович Р., Саблин М.В., Ларсон Г., Ханни К., Hitte C, Vigne JD Собаки сопровождали людей во время неолитической экспансии в Европу. Письма по биологии. 2018;14(10) [бесплатная статья PMC] [PubMed] [Google Scholar]44. Острандер Э.А., Уэйн Р.К., Фридман А.Х., Дэвис Б.В. Демографическая история, селекция и функциональное разнообразие собачьего генома.Природа Обзоры Генетика. 2017;18(12):705–720. doi: 10.1038/nrg.2017.67. [PubMed] [CrossRef] [Google Scholar]45. Pang JF, Kluetsch C, Zou XJ, Zhang AB, Luo LY, Angleby H, Ardalan A, Ekström C, Sköllermo A, Lundeberg J, Matsumura S, Leitner T, Zhang YP, Savolainen P данные мтДНК указывают на единое происхождение собак на юге реки Янцзы менее 16 300 лет назад от многочисленных волков.
В., Видга С., Виллерслев Э., Питулько В.В., Барнс И., Гилберт МТП, Добни К.М., Малхи Р.С., Мерчисон Э.П., Ларсон G, Frantz LAF Эволюционная история собак в Америке.Наука. 2018;361(6397):81–85. doi: 10.1126/science.aao4776. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]43. Оливье М., Трессе А., Франц Лаф, Брехард С., Бэлашеску А., Машкур М., Боронеант А., Пионье-Капитан М., Лебрассер О., Арбогаст Р.М., Бартосевич Л., Дебю К., Рабинович Р., Саблин М.В., Ларсон Г., Ханни К., Hitte C, Vigne JD Собаки сопровождали людей во время неолитической экспансии в Европу. Письма по биологии. 2018;14(10) [бесплатная статья PMC] [PubMed] [Google Scholar]44. Острандер Э.А., Уэйн Р.К., Фридман А.Х., Дэвис Б.В. Демографическая история, селекция и функциональное разнообразие собачьего генома.Природа Обзоры Генетика. 2017;18(12):705–720. doi: 10.1038/nrg.2017.67. [PubMed] [CrossRef] [Google Scholar]45. Pang JF, Kluetsch C, Zou XJ, Zhang AB, Luo LY, Angleby H, Ardalan A, Ekström C, Sköllermo A, Lundeberg J, Matsumura S, Leitner T, Zhang YP, Savolainen P данные мтДНК указывают на единое происхождение собак на юге реки Янцзы менее 16 300 лет назад от многочисленных волков. Молекулярная биология и эволюция. 2009;26(12):2849–2864. doi: 10.1093/molbev/msp195. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]46.Паркер Х.Г., ФонХолдт Б.М., Куиньон П., Маргулис Э.Х., Шао С., Мошер Д.С., Спади Т.К., Элькалун А., Каргилл М., Джонс П.Г., Маслен К.Л., Акланд Г.М., Саттер Н.Б., Куроки К., Бустаманте К.Д., Уэйн Р.К., Острандер Э.А. Экспрессированный ретроген Fgf4 связан с определяющей породу хондродисплазией у домашних собак. Наука. 2009;325(5943):995–998. doi: 10.1126/science.1173275. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]47. Plassais J, Rimbault M, Williams FJ, Davis BW, Schoenebeck JJ, Ostrander EA Анализ крупных и мелких собак выявил три гена на Х-хромосоме собак, связанные с массой тела, мускулатурой и толщиной спинного жира.Генетика PLoS. 2017;13(3):e1006661. doi: 10.1371/journal.pgen.1006661. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]48. Rimbault M, Beale HC, Schoenebeck JJ, Hoopes BC, Allen JJ, Kilroy-Glynn P, Wayne RK, Sutter NB, Ostrander EA Производные варианты шести генов объясняют почти половину уменьшения размера у пород собак.
Молекулярная биология и эволюция. 2009;26(12):2849–2864. doi: 10.1093/molbev/msp195. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]46.Паркер Х.Г., ФонХолдт Б.М., Куиньон П., Маргулис Э.Х., Шао С., Мошер Д.С., Спади Т.К., Элькалун А., Каргилл М., Джонс П.Г., Маслен К.Л., Акланд Г.М., Саттер Н.Б., Куроки К., Бустаманте К.Д., Уэйн Р.К., Острандер Э.А. Экспрессированный ретроген Fgf4 связан с определяющей породу хондродисплазией у домашних собак. Наука. 2009;325(5943):995–998. doi: 10.1126/science.1173275. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]47. Plassais J, Rimbault M, Williams FJ, Davis BW, Schoenebeck JJ, Ostrander EA Анализ крупных и мелких собак выявил три гена на Х-хромосоме собак, связанные с массой тела, мускулатурой и толщиной спинного жира.Генетика PLoS. 2017;13(3):e1006661. doi: 10.1371/journal.pgen.1006661. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]48. Rimbault M, Beale HC, Schoenebeck JJ, Hoopes BC, Allen JJ, Kilroy-Glynn P, Wayne RK, Sutter NB, Ostrander EA Производные варианты шести генов объясняют почти половину уменьшения размера у пород собак. Геномные исследования. 2013;23(12):1985–1995. doi: 10.1101/gr.157339.113. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]49. Сэтре П., Линдберг Дж., Леонард Дж. А., Олссон К., Петтерссон У., Эллегрен Х., Бергстрем Т.Ф., Вила С., Джазин Э. От дикого волка к домашней собаке: изменения экспрессии генов в мозге.Молекулярные исследования мозга. 2004;126(2):198–206. doi: 10.1016/j.molbrainres.2004.05.003. [PubMed] [CrossRef] [Google Scholar]50. Saetre P, Strandberg E, Sundgren PE, Pettersson U, Jazin E, Bergström TF Генетический вклад в формирование личности собак. Гены мозга и поведения. 2006;5(3):240–248. doi: 10.1111/j.1601-183X.2005.00155.x. [PubMed] [CrossRef] [Google Scholar]51. Саволайнен П., Лейтнер Т., Уилтон А.Н., Матису-Смит Э., Лундеберг Дж. Подробная картина происхождения австралийского динго, полученная в результате изучения митохондриальной ДНК.Труды Национальной академии наук Соединенных Штатов Америки. 2004;101(33):12387–12390. doi: 10.1073/pnas.0401814101. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]52.
Геномные исследования. 2013;23(12):1985–1995. doi: 10.1101/gr.157339.113. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]49. Сэтре П., Линдберг Дж., Леонард Дж. А., Олссон К., Петтерссон У., Эллегрен Х., Бергстрем Т.Ф., Вила С., Джазин Э. От дикого волка к домашней собаке: изменения экспрессии генов в мозге.Молекулярные исследования мозга. 2004;126(2):198–206. doi: 10.1016/j.molbrainres.2004.05.003. [PubMed] [CrossRef] [Google Scholar]50. Saetre P, Strandberg E, Sundgren PE, Pettersson U, Jazin E, Bergström TF Генетический вклад в формирование личности собак. Гены мозга и поведения. 2006;5(3):240–248. doi: 10.1111/j.1601-183X.2005.00155.x. [PubMed] [CrossRef] [Google Scholar]51. Саволайнен П., Лейтнер Т., Уилтон А.Н., Матису-Смит Э., Лундеберг Дж. Подробная картина происхождения австралийского динго, полученная в результате изучения митохондриальной ДНК.Труды Национальной академии наук Соединенных Штатов Америки. 2004;101(33):12387–12390. doi: 10.1073/pnas.0401814101. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]52. Саволайнен П., Чжан Ю.П., Луо Дж., Лундеберг Дж., Лейтнер Т. Генетические доказательства восточноазиатского происхождения домашних собак. Наука. 2002;298(5598):1610–1613. doi: 10.1126/science.1073906. [PubMed] [CrossRef] [Google Scholar]53. Шмутц С.М., Берриер Т.Г. Гены, влияющие на цвет и рисунок шерсти домашних собак: обзор.Генетика животных. 2007;38(6):539–549. doi: 10.1111/j.1365-2052.2007.01664.x. [PubMed] [CrossRef] [Google Scholar]54. Schmutz SM, Berryere TG, Goldfinch AD Генотипы TYRP1 и MC1R и их влияние на окрас шерсти у собак. Геном млекопитающих. 2002;13(7):380–387. doi: 10.1007/s00335-001-2147-2. [PubMed] [CrossRef] [Google Scholar]55. Шонебек Дж.Дж., Хатчинсон С.А., Байерс А., Бил Х.К., Кэррингтон Б., Фаден Д.Л., Римбо М., Декер Б., Кидд Дж.М., Суд Р., Бойко А.Р., Фондон Д.В., 3-й, Уэйн Р.К., Бустаманте К.Д., Цируна Б., Острандер Э.А. Вариация BMP3 способствует разнообразию черепа породы собак.Генетика PLoS. 2012;8(8):e1002849. doi: 10.1371/journal.pgen.1002849. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]57. Шеарин А.Л., Острандер Е.А. Впереди: собачьи модели геномики и болезней. Модели заболеваний и механизмы. 2010b;3(1−2):27–34. doi: 10.1242/dmm.004358. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]58. Spady TC, Ostrander EA Геномика псовых: картирование генов поведения чернобурой лисицы. Геномные исследования. 2007;17(3):259–263. doi: 10.1101/gr.6055807. [PubMed] [CrossRef] [Google Scholar]59.Саттер Н.Б., Бустаманте К.Д., Чейз К., Грей М.М., Чжао К.И., Чжу Л., Падукасахашрам Б., Карлинс Э., Дэвис С., Джонс П.Г., Куиньон П., Джонсон Г.С., Паркер Х.Г., Фретвелл Н., Мошер Д.С., Лоулер Д.Ф., Сатьярадж Э. , Nordborg M, Lark KG, Wayne RK, Ostrander EA Единственный аллель IGF1 является основной детерминантой маленького размера у собак. Наука. 2007;316(5821):112–115. doi: 10.1126/science.1137045. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]60. Tang B, Zhou Q, Dong L, Li W, Zhang X, Lan L, Zhai S, Xiao J, Zhang Z, Bao Y, Zhang YP, Wang GD, Zhao W iDog: интегрированный ресурс для домашних собак и диких псовых.
Саволайнен П., Чжан Ю.П., Луо Дж., Лундеберг Дж., Лейтнер Т. Генетические доказательства восточноазиатского происхождения домашних собак. Наука. 2002;298(5598):1610–1613. doi: 10.1126/science.1073906. [PubMed] [CrossRef] [Google Scholar]53. Шмутц С.М., Берриер Т.Г. Гены, влияющие на цвет и рисунок шерсти домашних собак: обзор.Генетика животных. 2007;38(6):539–549. doi: 10.1111/j.1365-2052.2007.01664.x. [PubMed] [CrossRef] [Google Scholar]54. Schmutz SM, Berryere TG, Goldfinch AD Генотипы TYRP1 и MC1R и их влияние на окрас шерсти у собак. Геном млекопитающих. 2002;13(7):380–387. doi: 10.1007/s00335-001-2147-2. [PubMed] [CrossRef] [Google Scholar]55. Шонебек Дж.Дж., Хатчинсон С.А., Байерс А., Бил Х.К., Кэррингтон Б., Фаден Д.Л., Римбо М., Декер Б., Кидд Дж.М., Суд Р., Бойко А.Р., Фондон Д.В., 3-й, Уэйн Р.К., Бустаманте К.Д., Цируна Б., Острандер Э.А. Вариация BMP3 способствует разнообразию черепа породы собак.Генетика PLoS. 2012;8(8):e1002849. doi: 10.1371/journal.pgen.1002849. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]57. Шеарин А.Л., Острандер Е.А. Впереди: собачьи модели геномики и болезней. Модели заболеваний и механизмы. 2010b;3(1−2):27–34. doi: 10.1242/dmm.004358. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]58. Spady TC, Ostrander EA Геномика псовых: картирование генов поведения чернобурой лисицы. Геномные исследования. 2007;17(3):259–263. doi: 10.1101/gr.6055807. [PubMed] [CrossRef] [Google Scholar]59.Саттер Н.Б., Бустаманте К.Д., Чейз К., Грей М.М., Чжао К.И., Чжу Л., Падукасахашрам Б., Карлинс Э., Дэвис С., Джонс П.Г., Куиньон П., Джонсон Г.С., Паркер Х.Г., Фретвелл Н., Мошер Д.С., Лоулер Д.Ф., Сатьярадж Э. , Nordborg M, Lark KG, Wayne RK, Ostrander EA Единственный аллель IGF1 является основной детерминантой маленького размера у собак. Наука. 2007;316(5821):112–115. doi: 10.1126/science.1137045. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]60. Tang B, Zhou Q, Dong L, Li W, Zhang X, Lan L, Zhai S, Xiao J, Zhang Z, Bao Y, Zhang YP, Wang GD, Zhao W iDog: интегрированный ресурс для домашних собак и диких псовых. Исследование нуклеиновых кислот. 2019;47(D1):D793–D800. doi: 10.1093/nar/gky1041. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]61. Танг Р., Нох Х.Дж., Ван Д., Сигурдссон С., Суоффорд Р., Перлоски М., Даксбери М., Паттерсон Э.Е., Олбрайт Дж., Кастельхано М., Аутон А., Бойко А.Р., Фенг Г., Линдблад-Тох К., Карлссон Е.К. Гены-кандидаты и функциональные некодирующие варианты, идентифицированные в собачьей модели обсессивно-компульсивного расстройства. Геномная биология. 2014;15(3):R25. doi: 10.1186/gb-2014-15-3-r25. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]62.Thawornwattana Y, Dalquen D, Yang Z Коалесцентный анализ филогеномных данных уверенно разрешает отношения между видами в комплексе видов anopheles gambiae. Молекулярная биология и эволюция. 2018;35(10):2512–2527. doi: 10.1093/molbev/msy158. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]63. Вила К., Саволайнен П., Мальдонадо Дж. Э., Аморим И. Р., Райс Дж. Э., Ханикатт Р. Л., Крэндалл К. А.
Исследование нуклеиновых кислот. 2019;47(D1):D793–D800. doi: 10.1093/nar/gky1041. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]61. Танг Р., Нох Х.Дж., Ван Д., Сигурдссон С., Суоффорд Р., Перлоски М., Даксбери М., Паттерсон Э.Е., Олбрайт Дж., Кастельхано М., Аутон А., Бойко А.Р., Фенг Г., Линдблад-Тох К., Карлссон Е.К. Гены-кандидаты и функциональные некодирующие варианты, идентифицированные в собачьей модели обсессивно-компульсивного расстройства. Геномная биология. 2014;15(3):R25. doi: 10.1186/gb-2014-15-3-r25. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]62.Thawornwattana Y, Dalquen D, Yang Z Коалесцентный анализ филогеномных данных уверенно разрешает отношения между видами в комплексе видов anopheles gambiae. Молекулярная биология и эволюция. 2018;35(10):2512–2527. doi: 10.1093/molbev/msy158. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]63. Вила К., Саволайнен П., Мальдонадо Дж. Э., Аморим И. Р., Райс Дж. Э., Ханикатт Р. Л., Крэндалл К. А. , Лундеберг Дж., Уэйн Р. К. Многочисленные и древние истоки домашней собаки. Наука. 1997;276(5319):1687–1689. дои: 10.1126/наука.276.5319.1687. [PubMed] [CrossRef] [Google Scholar]64. Вонхольдт Б.М., Поллинджер Дж.П., Ломюллер К.Е., Хан Э., Паркер Х.Г., Куиньон П., Дегенхардт Дж.Д., Бойко А.Р., Эрл Д.А., Аутон А., Рейнольдс А., Брайк К., Брисбин А., Ноулз Дж.К., Мошер Д.С., Спади Т.К., Элькалун А. , Геффен Э., Пилот М., Енджеевски В., Греко К., Рэнди Э., Баннаш Д., Уилтон А., Ширман Дж., Мусиани М., Каргилл М., Джонс П.Г., Цянь З., Хуанг В., Дин З.Л., Чжан Ю.П., Бустаманте К.Д., Острандер EA, Novembre J, Wayne RK Анализ SNP и гаплотипов всего генома раскрывает богатую историю, лежащую в основе одомашнивания собак.Природа. 2010;464(7290):898–902. doi: 10.1038/nature08837. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]65. Wang GD, Fan RX, Zhai W, Liu F, Wang L, Zhong L, Wu H, Yang HC, Wu SF, Zhu CL, Li Y, Gao Y, Ge RL, Wu CI, Zhang YP Генетическая конвергенция в адаптации собак и людей в высокогорную среду Тибетского нагорья.
, Лундеберг Дж., Уэйн Р. К. Многочисленные и древние истоки домашней собаки. Наука. 1997;276(5319):1687–1689. дои: 10.1126/наука.276.5319.1687. [PubMed] [CrossRef] [Google Scholar]64. Вонхольдт Б.М., Поллинджер Дж.П., Ломюллер К.Е., Хан Э., Паркер Х.Г., Куиньон П., Дегенхардт Дж.Д., Бойко А.Р., Эрл Д.А., Аутон А., Рейнольдс А., Брайк К., Брисбин А., Ноулз Дж.К., Мошер Д.С., Спади Т.К., Элькалун А. , Геффен Э., Пилот М., Енджеевски В., Греко К., Рэнди Э., Баннаш Д., Уилтон А., Ширман Дж., Мусиани М., Каргилл М., Джонс П.Г., Цянь З., Хуанг В., Дин З.Л., Чжан Ю.П., Бустаманте К.Д., Острандер EA, Novembre J, Wayne RK Анализ SNP и гаплотипов всего генома раскрывает богатую историю, лежащую в основе одомашнивания собак.Природа. 2010;464(7290):898–902. doi: 10.1038/nature08837. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]65. Wang GD, Fan RX, Zhai W, Liu F, Wang L, Zhong L, Wu H, Yang HC, Wu SF, Zhu CL, Li Y, Gao Y, Ge RL, Wu CI, Zhang YP Генетическая конвергенция в адаптации собак и людей в высокогорную среду Тибетского нагорья. Геномная биология и эволюция. 2014;6(8):2122–2128. doi: 10.1093/gbe/evu162. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]66. Wang GD, Zhai W, Yang HC, Wang L, Zhong L, Liu YH, Fan RX, Yin TT, Zhu CL, Poyarkov AD, Irwin DM, Hytönen MK, Lohi H, Wu CI, Savolainen P, Zhang YP Из южных Восточная Азия: естественная история домашних собак по всему миру.Клеточные исследования. 2016;26(1):21–33. doi: 10.1038/cr.2015.147. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]67. Wang GD, Zhai WW, Yang HC, Fan RX, Cao X, Zhong L, Wang L, Liu F, Wu H, Cheng LG, Poyarkov AD, Poyarkov NA, Tang SS, Zhao WM, Gao Y, Lv XM, Irwin DM , Саволайнен П., Ву К.И., Чжан Ю.П. Геномика отбора у собак и параллельная эволюция между собаками и людьми. Связь с природой. 2013; 4:1860. [PubMed] [Google Scholar]68. Уэбб А.А., Каллен С.Л. Цвет шерсти и неврологические и нейроофтальмологические заболевания, связанные с окраской шерсти.Канадский ветеринарный журнал. 2010;51(6):653–657. [Бесплатная статья PMC] [PubMed] [Google Scholar]69.
Геномная биология и эволюция. 2014;6(8):2122–2128. doi: 10.1093/gbe/evu162. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]66. Wang GD, Zhai W, Yang HC, Wang L, Zhong L, Liu YH, Fan RX, Yin TT, Zhu CL, Poyarkov AD, Irwin DM, Hytönen MK, Lohi H, Wu CI, Savolainen P, Zhang YP Из южных Восточная Азия: естественная история домашних собак по всему миру.Клеточные исследования. 2016;26(1):21–33. doi: 10.1038/cr.2015.147. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]67. Wang GD, Zhai WW, Yang HC, Fan RX, Cao X, Zhong L, Wang L, Liu F, Wu H, Cheng LG, Poyarkov AD, Poyarkov NA, Tang SS, Zhao WM, Gao Y, Lv XM, Irwin DM , Саволайнен П., Ву К.И., Чжан Ю.П. Геномика отбора у собак и параллельная эволюция между собаками и людьми. Связь с природой. 2013; 4:1860. [PubMed] [Google Scholar]68. Уэбб А.А., Каллен С.Л. Цвет шерсти и неврологические и нейроофтальмологические заболевания, связанные с окраской шерсти.Канадский ветеринарный журнал. 2010;51(6):653–657. [Бесплатная статья PMC] [PubMed] [Google Scholar]69. Вудс К.А., Камачо-Хюбнер С., Бартер Д., Кларк А.Дж., Сэвидж М.О. Делеция гена инсулиноподобного фактора роста I, вызывающая задержку внутриутробного развития и тяжелую низкорослость. Приложение Acta Paediatrica. 1997;86(S423):39–45. doi: 10.1111/j.1651-2227.1997.tb18367.x. [PubMed] [CrossRef] [Google Scholar]70. Вудс К.А., Камачо-Хюбнер С., Сэвидж М.О., Кларк А.Дж. Внутриутробная задержка роста и постнатальная недостаточность роста, связанные с делецией гена инсулиноподобного фактора роста I.Медицинский журнал Новой Англии. 1996;335(18):1363–1367. doi: 10.1056/NEJM199610313351805. [PubMed] [CrossRef] [Google Scholar]71. Wu DD, Ding XD, Wang S, Wójcik JM, Zhang Y, Tokarska M, Li Y, Wang MS, Faruque O, Nielsen R, Zhang Q, Zhang YP Повсеместная интрогрессия способствовала одомашниванию и адаптации в комплексе видов Bos . Экология природы и эволюция. 2018;2(7):1139–1145. [PubMed] [Google Scholar]72. Wu H, Liu YH, Wang GD, Yang CT, Otecko NO, Liu F, Wu SF, Wang L, Yu L, Zhang YP Идентификация молекулярных признаков адаптации к гипоксии по половым хромосомам: случай тибетского мастифа на основе анализа Х-хромосомы .
Вудс К.А., Камачо-Хюбнер С., Бартер Д., Кларк А.Дж., Сэвидж М.О. Делеция гена инсулиноподобного фактора роста I, вызывающая задержку внутриутробного развития и тяжелую низкорослость. Приложение Acta Paediatrica. 1997;86(S423):39–45. doi: 10.1111/j.1651-2227.1997.tb18367.x. [PubMed] [CrossRef] [Google Scholar]70. Вудс К.А., Камачо-Хюбнер С., Сэвидж М.О., Кларк А.Дж. Внутриутробная задержка роста и постнатальная недостаточность роста, связанные с делецией гена инсулиноподобного фактора роста I.Медицинский журнал Новой Англии. 1996;335(18):1363–1367. doi: 10.1056/NEJM199610313351805. [PubMed] [CrossRef] [Google Scholar]71. Wu DD, Ding XD, Wang S, Wójcik JM, Zhang Y, Tokarska M, Li Y, Wang MS, Faruque O, Nielsen R, Zhang Q, Zhang YP Повсеместная интрогрессия способствовала одомашниванию и адаптации в комплексе видов Bos . Экология природы и эволюция. 2018;2(7):1139–1145. [PubMed] [Google Scholar]72. Wu H, Liu YH, Wang GD, Yang CT, Otecko NO, Liu F, Wu SF, Wang L, Yu L, Zhang YP Идентификация молекулярных признаков адаптации к гипоксии по половым хромосомам: случай тибетского мастифа на основе анализа Х-хромосомы . Научные отчеты. 2016;6:35004. doi: 10.1038/srep35004. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Научные отчеты. 2016;6:35004. doi: 10.1038/srep35004. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]Окрас собак, объясненный модульными промоутерами древнего собачьего происхождения
Barsh, G., Gunn, T., He, L., Schlossman, S. & Дюк-Коэн, Дж. Биохимические и генетические исследования переключения типа пигмента. Пигментная ячейка Res. 13 , 48–53 (2000).
ПабМед Статья Google Scholar
Каро Т.и Малларино, Р. Окраска млекопитающих. Тренды Экол. Эвол. 35 , 357–366 (2020).
ПабМед Статья Google Scholar
Linnen, C.R. et al. Адаптивная эволюция нескольких признаков посредством множественных мутаций в одном гене. Наука 339 , 1312–1316 (2013).
КАС пабмед Статья Google Scholar
:format(png)/f906b0fead06ce1.ru.s.siteapi.org/img/b48eb87e30d2602144567e40b20a7486ca740cbc.jpg) «>
«>Врилинг, Х., Duhl, D.M., Millar, S.E., Miller, K.A. и Barsh, G.S. Различия в дорсальной и вентральной пигментации являются результатом региональной экспрессии гена агути мыши. Проц. Натл акад. науч. США 91 , 5667–5671 (1994).
КАС пабмед ПабМед Центральный Статья Google Scholar
Дрегер, Д. Л. и др. Атипичные генотипы сигнального белка собачьего агути предполагают новую хромосомную перестройку. Гены https://doi.org/10.3390/genes11070739 (2020).
Келин, С. Б. и Барш, Г. С. Генетика пигментации у собак и кошек. год. Преподобный Аним. Бионауч. 1 , 125–156 (2013).
ПабМед Статья КАС Google Scholar
Ритланд, К., Ньютон, К. и Маршалл, Х.Д. Наследование и структура популяции черного медведя «Кермод» с белой фазой. Курс. биол. 11 , 1468–1472 (2001).
биол. 11 , 1468–1472 (2001).
КАС пабмед Статья Google Scholar
Jones, M. R. et al. Адаптивная интрогрессия лежит в основе полиморфного сезонного камуфляжа зайцев-снегоступов. Наука 360 , 1355–1358 (2018).
КАС пабмед Статья Google Scholar
Гопалакришнан, С. и др. Межвидовой поток генов сформировал эволюцию рода Canis . Курс. биол. 28 , 3441–3449 (2018).
КАС пабмед ПабМед Центральный Статья Google Scholar
Koepfli, K.P. et al. Полногеномные данные показывают, что африканские и евразийские золотые шакалы являются разными видами. Курс. биол. 25 , 2158–2165 (2015).
КАС пабмед Статья Google Scholar
 «>
«>Ван М.С. и др. Древняя гибридизация с неизвестной популяцией способствовала адаптации псовых к высокогорью. Мол. биол. Эвол . https://doi.org/10.1093/molbev/msaa113 (2020 г.).
Gipson, P. S. et al. Цветовые узоры Хименеса среди волков в западной части Северной Америки. Дикий. соц. Бык. 30 , 821–830 (2002).
Google Scholar
Zhang, W. et al. Адаптации к гипоксии у серого волка ( Canis lupus chanco ) с Цинхай-Тибетского нагорья. Генетика PLoS. 10 , e1004466 (2014).
ПабМед ПабМед Центральный Статья КАС Google Scholar
Райт С. Наследование цвета у млекопитающих. Дж. Херед. 8 , 224–235 (1917).
Артикул Google Scholar
Литтл, CC Наследование цвета шерсти у собак (Comstock Publishing Associates, 1957).
Дрегер Д. Л., Паркер Х. Г., Острандер Э. А. и Шмутц С. М. Идентификация мутации, связанной с фенотипами седло-подпалого и черно-подпалого у бассет-хаундов и вельш-корги пемброк. Дж. Херед. 104 , 399–406 (2013).
КАС пабмед Статья Google Scholar
Берриер, Т. Г., Кернс, Дж. А., Барш, Г. С. и Шмутц, С. М. Ассоциация аллеля агути с палевым или соболиным окрасом шерсти у домашних собак. Мамм. Геном 16 , 262–272 (2005).
КАС пабмед Статья Google Scholar
Giska, I. et al. Интрогрессия вызывает повторную эволюцию полиморфизма окраски зимнего окраса у зайцев. Проц. Натл акад. науч. США 116 , 24150–24156 (2019).
КАС пабмед ПабМед Центральный Статья Google Scholar
 «>
«>Баиз М.Д., Вуд, А.В., Брелсфорд, А., Ловетт, И.Дж. и Тэйвс, Д.П.Л. Гены пигментации свидетельствуют о повторной дивергенции и множественных приступах интрогрессии у камышевок Setophaga . Курс. биол. 31 , 643–649 (2021).
КАС пабмед Статья Google Scholar
Kim, K.W. et al. Генетика и доказательства уравновешивающего выбора сцепленного с полом полиморфизма окраски у певчих птиц. Нац. коммун. 10 , 1852 (2019).
ПабМед ПабМед Центральный Статья КАС Google Scholar
Toews, D. P. et al. Гены оперения и мало что отличает геномы гибридизирующихся славок. Курс. биол. 26 , 2313–2318 (2016).
КАС пабмед Статья Google Scholar
Ван С.и другие. Отбор на небольшой области генома лежит в основе дифференциации по множеству цветовых признаков между двумя видами камышевок. Эволюция. лат. 4 , 502–515 (2020).
Эволюция. лат. 4 , 502–515 (2020).
ПабМед ПабМед Центральный Статья Google Scholar
Henkel, J. et al. Сигнатуры селекции у коз выявляют варианты числа копий, лежащие в основе фенотипов окраски шерсти, определяющих породу. Генетика PLoS. 15 , e1008536 (2019).
КАС пабмед ПабМед Центральный Статья Google Scholar
Fontanesi, L. et al. Характеристика гена сигнального белка агути кролика (ASIP): транскрипты, филогенетический анализ и идентификация причинной мутации черного окраса шерсти нонагути. Геномика 95 , 166–175 (2010).
КАС пабмед Статья Google Scholar
Летко А.и другие. Делеция, охватывающая промотор и первый экзон специфичной для цикла волос изоформы транскрипта ASIP у черно-подпалых кроликов. Аним. Жене. 51 , 137–140 (2020).
Аним. Жене. 51 , 137–140 (2020).
КАС пабмед Статья Google Scholar
Дуль, Д. М., Врилинг, Х., Миллер, К. А., Вольф, Г. Л. и Барш, Г. С. Неоморфные мутации агути у желтых мышей с ожирением. Нац. Жене. 8 , 59–65 (1994).
КАС пабмед Статья Google Scholar
Бальм, Дж., О’Коннор, С. и Фэллон, С. Новые даты на костях динго из пещеры Мадура являются старейшими убедительными доказательствами прибытия этого вида в Австралию. науч. Респ. 8 , 9933 (2018).
ПабМед ПабМед Центральный Статья КАС Google Scholar
Lee, E.J. et al.Анализ древней ДНК древнейших видов псовых из сибирской Арктики и генетический вклад в развитие домашней собаки. PLoS ONE 10 , e0125759 (2015).
ПабМед ПабМед Центральный Статья КАС Google Scholar
Синдинг, М.-Х. С. и др. Адаптированные к Арктике собаки появились на переходе от плейстоцена к голоцену. Наука 368 , 1495–1499 (2020).
КАС пабмед ПабМед Центральный Статья Google Scholar
Вентилятор, З.и другие. Мировые закономерности геномной изменчивости и примесей у серых волков. Рез. генома. 26 , 163–173 (2016).
КАС пабмед ПабМед Центральный Статья Google Scholar
Луг, Л. и др. Древняя ДНК предполагает, что современные волки ведут свое происхождение от экспансии позднего плейстоцена из Берингии. Мол. Экол . https://doi.org/10.1111/mec.15329 (2019 г.).
Дрегер Д. Л. и Шмутц, С. М. Вставка SINE вызывает фенотипы черно-подпалого и седло-подпалого у домашних собак. Дж. Херед. 102 , С11–С18 (2011).
Л. и Шмутц, С. М. Вставка SINE вызывает фенотипы черно-подпалого и седло-подпалого у домашних собак. Дж. Херед. 102 , С11–С18 (2011).
КАС пабмед Статья Google Scholar
Фридман А. Х. и Уэйн Р. К. Расшифровка происхождения собак: от окаменелостей к геномам. год. Преподобный Аним. Бионауч. 5 , 281–307 (2017).
ПабМед Статья Google Scholar
Добин А.и другие. STAR: сверхбыстрый универсальный выравниватель RNA-seq. Биоинформатика 29 , 15–21 (2013).
КАС пабмед ПабМед Центральный Статья Google Scholar
Robinson, J. T. et al. Интегративный просмотрщик геномики. Нац. Биотехнолог. 29 , 24–26 (2011).
КАС пабмед ПабМед Центральный Статья Google Scholar
 «>
«>Джаганнатхан, В., Drogemuller, C. & Leeb, T. Консорциум базы данных биомедицинских вариантов собак Полный каталог биомедицинских вариантов, основанный на последовательностях полных геномов 582 собак и восьми волков. Аним. Жене. 50 , 695–704 (2019).
КАС пабмед ПабМед Центральный Статья Google Scholar
RepeatMasker Open v.4.0 (Институт системной биологии, 2013 г.).
Ислам, С. и др. Характеристика транскрипционного ландшафта одиночных клеток с помощью высокомультиплексной РНК-секвенции. Рез. генома. 21 , 1160–1167 (2011).
КАС пабмед ПабМед Центральный Статья Google Scholar
Крюцков К. и др. Снижение мРНК глобина для секвенирования транскриптома цельной крови. науч. Респ. 6 , 31584 (2016).
КАС пабмед ПабМед Центральный Статья Google Scholar
 «>
«>Пертеа, М., Ким, Д., Пертеа, Г. М., Лик, Дж. Т. и Зальцберг, С. Л. Анализ экспрессии на уровне транскрипта в экспериментах по секвенированию РНК с помощью HISAT, StringTie и Ballgown. Нац. протокол 11 , 1650–1667 (2016).
КАС пабмед ПабМед Центральный Статья Google Scholar
Брей, Н. Л., Пиментел, Х., Мелстед, П. и Пачтер, Л. Почти оптимальная вероятностная количественная оценка секвенирования РНК. Нац. Биотехнолог. 34 , 525–527 (2016).
КАС пабмед Статья Google Scholar
Ли, Х. и Дурбин, Р. Быстрое и точное выравнивание коротких считываний с помощью преобразования Берроуза-Уилера. Биоинформатика 25 , 1754–1760 (2009).
КАС пабмед ПабМед Центральный Статья Google Scholar
 «>
«>Риммер, А. и др. Интеграция подходов на основе картирования, сборки и гаплотипов для вызова вариантов в приложениях для клинического секвенирования. Нац. Жене. 46 , 912–918 (2014).
КАС пабмед ПабМед Центральный Статья Google Scholar
Danecek, P. et al. Вариант формата вызова и VCFtools. Биоинформатика 27 , 2156–2158 (2011).
КАС пабмед ПабМед Центральный Статья Google Scholar
Браунинг, С. Р. и Браунинг, Б.L. Быстрое и точное определение фаз гаплотипов и вывод отсутствующих данных для полногеномных ассоциативных исследований с использованием локальной кластеризации гаплотипов. утра. Дж. Хам. Жене. 81 , 1084–1097 (2007).
КАС пабмед ПабМед Центральный Статья Google Scholar
 «>
«>Кумар, С., Стечер, Г., Ли, М., Княз, К. и Тамура, К. MEGA X: молекулярно-эволюционный генетический анализ на вычислительных платформах. Мол.биол. Эвол. 35 , 1547–1549 (2018).
КАС пабмед ПабМед Центральный Статья Google Scholar
Стечер Г., Тамура К. и Кумар С. Молекулярно-эволюционный генетический анализ (MEGA) для macOS. Мол. биол. Эвол. 37 , 1237–1239 (2020).
КАС пабмед ПабМед Центральный Статья Google Scholar
фон Холдт, Б.М. и др. Полногеномный взгляд на эволюционную историю загадочных волкоподобных псовых. Рез. генома. 21 , 1294–1305 (2011).
КАС пабмед ПабМед Центральный Статья Google Scholar
Botigué, L. R. et al. Геномы древних европейских собак обнаруживают преемственность со времен раннего неолита. Нац. коммун. 8 , 16082 (2017).
Нац. коммун. 8 , 16082 (2017).
ПабМед ПабМед Центральный Статья КАС Google Scholar
Франц, Л.А. и др. Геномные и археологические данные свидетельствуют о двойном происхождении домашних собак. Наука 352 , 1228–1231 (2016).
КАС пабмед Статья Google Scholar
Ni Leathlobhair, M. et al. Эволюционная история собак в Америке. Наука 361 , 81–85 (2018).
КАС пабмед Статья Google Scholar
Скоглунд, П., Ersmark, E., Palkopoulou, E. & Dalen, L. Древний геном волка показывает раннее расхождение предков домашних собак и примесь высокоширотных пород. Курс. биол. 25 , 1515–1519 (2015).
КАС пабмед Статья Google Scholar
 «>
«>vonHoldt, B.M. et al. Идентификация недавней гибридизации между серыми волками и домашними собаками с помощью генотипирования SNP. Мамм. Геном 24 , 80–88 (2013).
КАС пабмед Статья Google Scholar
ScienceShot: Расшифровка зевоты собаки | Наука
Страна: Страна * AfghanistanAland IslandsAlbaniaAlgeriaAndorraAngolaAnguillaAntarcticaAntigua и BarbudaArgentinaArmeniaArubaAustraliaAustriaAzerbaijanBahamasBahrainBangladeshBarbadosBelarusBelgiumBelizeBeninBermudaBhutanBolivia, многонациональное государство ofBonaire, Синт-Эстатиус и SabaBosnia и HerzegovinaBotswanaBouvet IslandBrazilBritish Индийский океан TerritoryBrunei DarussalamBulgariaBurkina FasoBurundiCambodiaCameroonCanadaCape VerdeCayman IslandsCentral африканских RepublicChadChileChinaChristmas IslandCocos (Килинг) IslandsColombiaComorosCongoCongo, Демократическая Республика theCook IslandsCosta RicaCote D’IvoireCroatiaCubaCuraçaoCyprusCzech RepublicDenmarkDjiboutiDominicaDominican RepublicEcuadorEgyptEl SalvadorEquatorial GuineaEritreaEstoniaEthiopiaFalkland острова (Мальвинские )Фарерские островаФиджиФинляндияФранцияФранцузская ГвианаФранцузская ПолинезияФранцузские Южные ТерриторииГабонГамбияГрузияГерманияГанаГибралтарГрецияГренландияГваделупаГватемалаГернсиГвинеяГвинея-БисауГайанаГаитиОстров Херд и МакДональда IslandsHoly Престол (Ватикан) HondurasHong KongHungaryIcelandIndiaIndonesiaIran, Исламская Республика ofIraqIrelandIsle из ManIsraelItalyJamaicaJapanJerseyJordanKazakhstanKenyaKiribatiKorea, Корейская Народно-Демократическая Республика ofKorea, Республика ofKuwaitKyrgyzstanLao Народная Демократическая RepublicLatviaLebanonLesothoLiberiaLibyan Арабская JamahiriyaLiechtensteinLithuaniaLuxembourgMacaoMacedonia, бывшая югославская Республика ofMadagascarMalawiMalaysiaMaldivesMaliMaltaMartiniqueMauritaniaMauritiusMayotteMexicoMoldova, Республика ofMonacoMongoliaMontenegroMontserratMoroccoMozambiqueMyanmarNamibiaNauruNepalNetherlandsNew CaledoniaNew ZealandNicaraguaNigerNigeriaNiueNorfolk IslandNorwayOmanPakistanPalestinianPanamaPapua Новый GuineaParaguayPeruPhilippinesPitcairnPolandPortugalQatarReunionRomaniaRussian FederationRWANDASaint BarthélemySaint Елены, Вознесения и Тристан-да-КуньяСент-Китс и НевисСент-ЛюсияСент-Мартен (французская часть)Сен-Пьер и МикелонСент-Винсент и ГренадиныСэм oaSan MarinoSao Том и PrincipeSaudi ArabiaSenegalSerbiaSeychellesSierra LeoneSingaporeSint Маартен (Голландская часть) SlovakiaSloveniaSolomon IslandsSomaliaSouth AfricaSouth Джорджия и Южные Сандвичевы IslandsSouth SudanSpainSri LankaSudanSurinameSvalbard и Ян MayenSwazilandSwedenSwitzerlandSyrian Arab RepublicTaiwanTajikistanTanzania, Объединенная Республика ofThailandTimor-LesteTogoTokelauTongaTrinidad и TobagoTunisiaTurkeyTurkmenistanTurks и Кайкос IslandsTuvaluUgandaUkraineUnited арабского EmiratesUnited KingdomUnited StatesUruguayUzbekistanVanuatuVenezuela, Боливарианская Республика ofVietnamVirgin остров, BritishWallis и ФутунаЗападная СахараЙеменЗамбияЗимбабве
Пожертвовать сейчасПоддержите некоммерческую научную журналистику
Если мы чему-то и научились во время пандемии COVID-19, так это тому, что мы не можем ждать реакции кризиса. Science и AAAS неустанно работают над предоставлением достоверной, основанной на фактических данных информации о последних научных исследованиях и политике с обширным бесплатным освещением пандемии. Ваш не облагаемый налогом вклад играет решающую роль в поддержании этих усилий.
Science и AAAS неустанно работают над предоставлением достоверной, основанной на фактических данных информации о последних научных исследованиях и политике с обширным бесплатным освещением пандемии. Ваш не облагаемый налогом вклад играет решающую роль в поддержании этих усилий.
Расшифровка снаряжения для собак — Загородный магазин — Ферма — Домашнее животное
Есть, спать, гулять, играть; день в жизни собаки довольно прост.Однако выбор правильного снаряжения для вашей собаки может быть довольно запутанным. В большинстве зоомагазинов представлены сотни марок, размеров и форм ошейников, игрушек и поводков. Прочтите несколько советов о том, как правильно выбрать снаряжение для вашей собаки.
Большинству собак требуется очень простое оборудование для кормления. Самая важная часть выбора мисок для еды и воды — убедиться, что они подходят по размеру для вашей собаки. Если у вас маленький щенок, подумайте о том, какого размера будет собака, когда вырастет, и соответственно подберите миски.Металлические и керамические миски считаются более гигиеничными, чем пластиковые миски, которые могут задерживать бактерии в царапинах и порезах. Подберите коврик для собачьих салфеток, чтобы собирать разливы и капли.
Самая важная часть выбора мисок для еды и воды — убедиться, что они подходят по размеру для вашей собаки. Если у вас маленький щенок, подумайте о том, какого размера будет собака, когда вырастет, и соответственно подберите миски.Металлические и керамические миски считаются более гигиеничными, чем пластиковые миски, которые могут задерживать бактерии в царапинах и порезах. Подберите коврик для собачьих салфеток, чтобы собирать разливы и капли.
Каждой собаке нужен прочный ошейник и поводок. Базовый нейлоновый или кожаный ошейник идеально подходит для повседневного использования. Если есть возможность, возьмите собаку с собой в зоомагазин, чтобы примерить стили и размеры ошейников перед покупкой. Если вы не можете принести свою собаку в зоомагазин, измерьте ее шею и добавьте несколько дюймов, чтобы найти правильный размер ошейника.
Воротник должен плотно прилегать к верхней части шеи. Он должен быть достаточно тугим, чтобы между шеей и воротником проходило всего два пальца. Он не должен соскальзывать через голову собаки или спускаться ей на плечи. Если у вас растущий щенок, не забывайте часто проверять ошейник, чтобы убедиться, что он не становится слишком тесным. Дрессировочные ошейники, которые душит, щиплют или точат собак, следует использовать только при активном обучении вашей собаки и никогда не оставлять на собаке постоянно.
Он не должен соскальзывать через голову собаки или спускаться ей на плечи. Если у вас растущий щенок, не забывайте часто проверять ошейник, чтобы убедиться, что он не становится слишком тесным. Дрессировочные ошейники, которые душит, щиплют или точат собак, следует использовать только при активном обучении вашей собаки и никогда не оставлять на собаке постоянно.
После того, как вы выбрали ошейник, найдите подходящий поводок.Ищите поводок с фурнитурой, которая соответствует размеру фурнитуры на ошейнике. Маленьким собакам и щенкам лучше всего подойдет легкий поводок. Большим собакам понадобится что-то более существенное. По мере роста собаки вам может понадобиться обновить ошейник и поводок.
Выдвижной поводок — хороший вариант, чтобы позволить вашей собаке исследовать территорию, оставаясь при этом под вашим контролем. Выдвижной поводок может не помочь научить вашу собаку правильным манерам ходьбы на поводке. Тем не менее, убирающиеся поводки могут быть полезны при посещении парков и общественных мест, которые вы хотите исследовать, но ваша собака должна оставаться на поводке.
Конура и лежанка пригодятся для приучения собаки к клетке. Собаки любят иметь собственное пространство для сна и отдыха. Лежаки и клетки для собак бывают самых разных форм, размеров и материалов. Ищите кровать и клетку, которые немного больше, чем ваша собака будет в натуральную величину. Поместите в клетку несколько игрушек, чтобы помочь собаке понять, что это ее пространство.
Игрушки — отличный способ пообщаться с собакой и дать ей занятие, когда вы не можете играть. Доступны сотни игрушек, но большинство собак будут довольны простым мячиком и канатом.Если ваша собака склонна грызть предметы, запаситесь костями и игрушками для жевания, чтобы отвлечь ее внимание от мебели и обуви.
Помните, что собаки любят бегать и играть на улице и, скорее всего, будут грубо обращаться с оборудованием. Если ваша собака уже взрослая, часто выгодно инвестировать в высококачественные продукты, которые выдержат испытание временем. В Country Store представлен ряд высококачественных брендов собачьего снаряжения. Если у вас есть вопросы о том, какие продукты лучше всего подходят для вашей собаки, просто зайдите и спросите!
Если у вас есть вопросы о том, какие продукты лучше всего подходят для вашей собаки, просто зайдите и спросите!
Объяснение концовки «Сила собаки»
Сила собаки создан знаменитым режиссером Джейн Кэмпион, которая была лишь второй женщиной, номинированной на премию Оскар за лучшую режиссуру в 1994 году за фильм Фортепиано .Она берет исходный материал из одноименного романа Томаса Сэвиджа. Человек, специализирующийся на вестернах на тему угнетения, вызванного токсичной мужественностью.
Здесь она снимает фильм, в котором главный герой Фил Бербанк (Бенедикт Камбербэтч) работает погонщиком скота и владеет ранчо вместе со своим братом Джорджем (Джесси Племонс). Джордж добр, сердечен и обладает способностью сопереживать, которой обладают немногие мужчины. Фил — это портрет мужской токсичности, потому что мы узнаем, что он скрытый гомосексуал из Монтаны 1920-х годов.Он играет в жестокую психологическую игру со своей невесткой Роуз (Кирстен Данст) и ее сыном Питером ( «Дорога», Коди Смит-Макфи), которого Фил считает геем, потому что он женоподобный. Он пытается взять его под свое крыло, как это сделал для него Бронко Генри.
Фильм Netflix Сила собаки Объяснение концовки
Когда Питер находит тайное убежище Фила, пещеру, сделанную из ветвей и корней деревьев, которые образуют естественный барьер и доступ к ручью, он обнаруживает рычаг.Питер обнаруживает, что у Фила есть коробка, полная непристойных журналов и вырезок, все с голыми мужчинами. Теперь ясно, что токсичная мужественность родилась из-за того, что его угнетало общество, которое не принимало его, и из-за того, что он использовал ковбойский образ жизни в качестве бородатого камуфляжа. Были ли у Питера интимные отношения с Бронко Генри, или он сделал из него мужчину, не мучающего из него «гея»?
Были ли у Питера интимные отношения с Бронко Генри, или он сделал из него мужчину, не мучающего из него «гея»?
В любом случае, Фил мучает свою мать. У него не может быть этого. Поэтому он играет в соблазнительную психологическую игру, и Питер дает ему попробовать свое собственное лекарство.Он позволяет Филу обучать его, чтобы наладить связь, в то время как Питер, который учится на врача, по неизвестным причинам препарирует мертвого быка. Когда Питер привлекает Фила дружбой и соблазнением, он позволяет ему затянуться сигаретой. У Фила одышка, лихорадка, озноб, головокружение, проливной пот и сильная тошнота через несколько часов. Он умирает через день или два.
Выяснилось, что Фил умер от отравления сибирской язвой. Но мы видели, что он был специалистом и очень осторожен в защите от сибирской язвы при обращении со скотом.Как Фил мог быть таким беспечным? Ну, это потому, что Питер дал ему кусок кожи, который он отрезал от крупного рогатого скота (я неправильно приписал его сигарете, которую он ему дал, так что даже ее использование отвлекло не только Фила, но и меня), пропитанную бактериальной болезнью. . Теперь, когда Фил умер, Роуз может счастливо жить с Джорджем (Джесси Племмонс) и продолжать свои медицинские исследования.
. Теперь, когда Фил умер, Роуз может счастливо жить с Джорджем (Джесси Племмонс) и продолжать свои медицинские исследования.
Сериал The Power of the Dog от Netflix представляет собой многослойный взгляд на двойственность нетерпимости, сексуальной ориентации и мужской токсичности.Финал демонстрирует твердое понимание того, что это увлекательное исследование персонажей о том, как социальное угнетение имеет непреднамеренные последствия превращения маргинализированных в угнетателей. Это вестерн о порочном круге, который никогда не заканчивается, и никто не знает, где он начался.
Что вы думаете о финале фильма Netflix «Сила собаки»? Комментарий ниже!
РодственныеВспышки чумы в популяциях луговых собачек, объясняемые порогами перколяции при множественности альтернативных хозяев грызунов (1).Чума не является историческим пережитком и остается актуальной проблемой общественного здравоохранения: недавно были вспышки среди населения (например, на Мадагаскаре, Уганде и Китае), несколько случаев заболевания людей происходят в Соединенных Штатах каждый год, а возбудитель представляет собой потенциальная угроза как агент биотерроризма (1–5).
 Завезенная в Соединенные Штаты примерно в 1900 году, чума в настоящее время представляет угрозу для сохранения нескольких видов животных, в том числе популяций чернохвостой луговой собачки ( Cynomys ludivicianus ), вида, вызывающего озабоченность при сохранении пастбищ Северной Америки (6).Луговые собачки — это социальные суслики, населяющие большие города или колонии в прериях, которые могут простираться на сотни гектаров и насчитывать несколько тысяч особей. Вызванная чумой гибель луговых собачек происходит периодически, но когда это происходит, почти 100% луговых собачек в колонии умирают от болезни (7, 8). Основываясь на наблюдениях за гибелью луговых собачек, общий консенсус предполагает, что вспышки чумы происходят чрезвычайно быстро, так что лишь несколько выживших наблюдаются через 6–8 недель (7, 9).Однако предполагаемая короткая продолжительность эпизоотии может быть предвзятой, потому что подтверждение чумы часто зависит от достаточной удачи для обнаружения туши луговых собачек над землей (большинство луговых собачек умирает в своих норах), а систематическое наблюдение за чумой проводится нечасто.
Завезенная в Соединенные Штаты примерно в 1900 году, чума в настоящее время представляет угрозу для сохранения нескольких видов животных, в том числе популяций чернохвостой луговой собачки ( Cynomys ludivicianus ), вида, вызывающего озабоченность при сохранении пастбищ Северной Америки (6).Луговые собачки — это социальные суслики, населяющие большие города или колонии в прериях, которые могут простираться на сотни гектаров и насчитывать несколько тысяч особей. Вызванная чумой гибель луговых собачек происходит периодически, но когда это происходит, почти 100% луговых собачек в колонии умирают от болезни (7, 8). Основываясь на наблюдениях за гибелью луговых собачек, общий консенсус предполагает, что вспышки чумы происходят чрезвычайно быстро, так что лишь несколько выживших наблюдаются через 6–8 недель (7, 9).Однако предполагаемая короткая продолжительность эпизоотии может быть предвзятой, потому что подтверждение чумы часто зависит от достаточной удачи для обнаружения туши луговых собачек над землей (большинство луговых собачек умирает в своих норах), а систематическое наблюдение за чумой проводится нечасто.
Примечательная картина чумы, состоящая из спорадических взрывоопасных вспышек (эпизоотическая/эпидемическая фаза), за которыми следуют длительные периоды скрытого затишья (энзоотическая/поддерживающая фаза), характерна для многих других смертельно опасных болезней человека (например,г., хантавирусы, вирус хендра и сибирская язва). Часто ключевым недостающим элементом усилий по контролю и искоренению таких вирулентных болезней является понимание того, где патогены сохраняются в энзоотические периоды, и способность идентифицировать условия, которые приводят к вспышкам или эпизоотиям. Существуют две общие гипотезы, объясняющие энзоото-эпизоотическую динамику. Первая гипотеза постулирует, что патогены переходят от одного вида хозяина к очень восприимчивому виду хозяина, который страдает высокой смертностью после заражения.Вторая гипотеза постулирует, что патогены просто циркулируют в пределах одного вида хозяина и что эпизоотии являются результатом повышенной скорости передачи, возможно, в результате изменений условий окружающей среды, численности хозяина/переносчика и/или поведения (1, 10). Из-за непредсказуемого характера спорадически возникающих смертельных заболеваний существует несколько полевых исследований свободноживущих популяций хозяев, которым удалось различить эти гипотезы (11).
Из-за непредсказуемого характера спорадически возникающих смертельных заболеваний существует несколько полевых исследований свободноживущих популяций хозяев, которым удалось различить эти гипотезы (11).
Высокая смертность и быстрое распространение чумы в популяциях луговых собачек привели к общему мнению, что возбудитель, скорее всего, отсутствует в здоровых колониях луговых собачек и вместо этого сохраняется во время межэпизоотий у альтернативных видов грызунов (например,g., оленьих мышей), хотя данных, подтверждающих этот механизм, немного (1, 9, 12–14). Чума также может сохраняться среди блох-переносчиков после того, как местные популяции млекопитающих-хозяев были истреблены, хотя блохи луговых собачек ( Oropsylla hirsuta ) характеризуются высокой смертностью при отсутствии питания кровью, а численность блох заметно снижается после эпизоотии (15, 16). Тем не менее, поскольку Y. pestis является универсальным патогеном, который может заражать несколько видов хозяев и блох в сообществе прерий, чума в колониях луговых собачек представляет собой идеальную систему для изучения гипотез распространения и персистенции одного хозяина для экологии энзоото-эпизоотических патогенов. .
.
Для изучения энзоотической персистенции и эпизоотических вспышек чумы мы смоделировали динамику Y. pestis в колониях луговых собачек, используя информацию из полевых данных, которые позволяют предположить, что северная мышь-кузнечик ( Onychomys leucogaster ) тесно связана с чумой. экология. Мыши-кузнечики заражаются чумой во время эпизоотии луговых собачек, но их смертность неодинакова. Высокая численность кузнечиковых мышей связана с более высокой вероятностью эпизоотий чумы в колониях луговых собачек, а во время эпизоотий кузнечиковые мыши содержат большое количество O.hirsuta , блоха луговых собачек, обычно зависящая от хозяина (17).
Результаты и обсуждение
В отсутствие кузнечиков чума имела тенденцию заражать лишь небольшую часть популяции луговых собачек в течение 5 лет (рис. 1). Хотя в некоторых случаях около 20% популяции луговых собачек были инфицированы в течение 5-летнего периода, ежедневные показатели заболеваемости были небольшими (редко превышали 6 случаев в день). Семьи луговых собачек защищают территорию своего кружка от соседних луговых собачек, но в случае гибели всех членов кружка луговые собачки обычно присоединяются к одной соседней семье (18).Следовательно, чума имела тенденцию последовательно передаваться между соседними группами, потому что передача инфицированных блох происходит, когда зараженная луговая собачка умирает и блохи спрыгивают с туши на другую луговую собачку. Другими словами, преобладающие пути передачи блохами от хозяина к хозяину из одной группы в другую сильно ограничены в пространстве, и, поскольку чума может распространяться от одной группы к другой только последовательно, эпизоотии не могут возникать только благодаря этому процессу. Даже с учетом расселения луговых собачек чума по-прежнему может перемещаться между соседними группами только после того, как она переместилась на новую семейную территорию, и, следовательно, эпизоотии остаются маловероятными.
Семьи луговых собачек защищают территорию своего кружка от соседних луговых собачек, но в случае гибели всех членов кружка луговые собачки обычно присоединяются к одной соседней семье (18).Следовательно, чума имела тенденцию последовательно передаваться между соседними группами, потому что передача инфицированных блох происходит, когда зараженная луговая собачка умирает и блохи спрыгивают с туши на другую луговую собачку. Другими словами, преобладающие пути передачи блохами от хозяина к хозяину из одной группы в другую сильно ограничены в пространстве, и, поскольку чума может распространяться от одной группы к другой только последовательно, эпизоотии не могут возникать только благодаря этому процессу. Даже с учетом расселения луговых собачек чума по-прежнему может перемещаться между соседними группами только после того, как она переместилась на новую семейную территорию, и, следовательно, эпизоотии остаются маловероятными.
Взаимосвязь между активностью чумы, плотностью кузнечиков p M и частотой прыжков блох j P , измеренной с помощью ( A ) общего числа инфицированных луговых собачек во время моделирования и ( B ) продолжительность активности чумы в днях.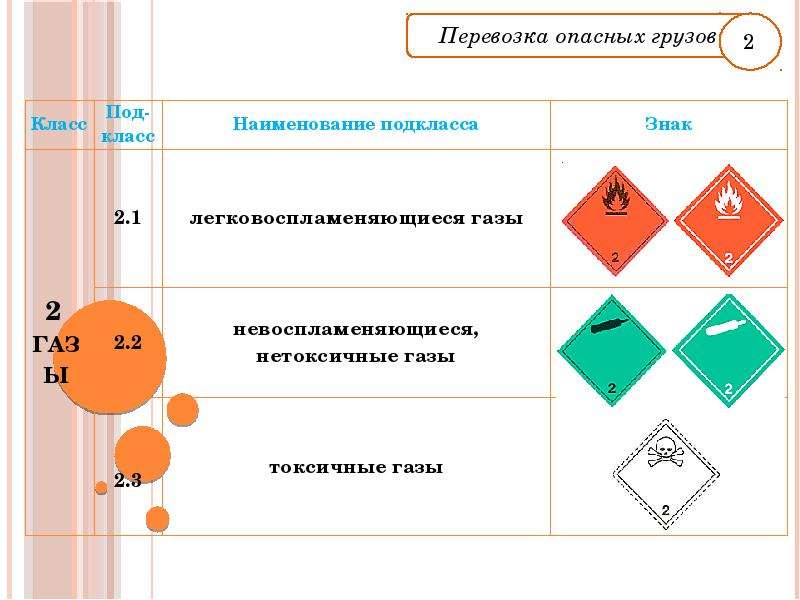 Каждый из 625 пикселей представляет собой среднее из 100 ( A и B ) или 1000 ( C и D ) запусков моделирования, кодированных цветом по значению (см. цветную полосу справа от каждого графика).Оси логарифмически масштабированы, хотя левый столбец каждой панели представляет 0 мышей. Эпизоотический порог хорошо виден у А . B показывает, что чума может сохраняться в популяциях луговых собачек в течение длительных периодов времени, несмотря на высокую летальность. При низкой плотности мышей вспышки быстро затухают и, следовательно, имеют короткую продолжительность. При средней плотности эпизоотии медленно прожигают колонию, тогда как при высокой плотности котерии настолько хорошо связаны, что чума быстро распространяется по колонии.( C ) Индивидуальный R 0 . Белый цвет обозначает значения <1. ( D ) На основе Coterie R 0 ( R *) с тем же цветовым кодом.
Каждый из 625 пикселей представляет собой среднее из 100 ( A и B ) или 1000 ( C и D ) запусков моделирования, кодированных цветом по значению (см. цветную полосу справа от каждого графика).Оси логарифмически масштабированы, хотя левый столбец каждой панели представляет 0 мышей. Эпизоотический порог хорошо виден у А . B показывает, что чума может сохраняться в популяциях луговых собачек в течение длительных периодов времени, несмотря на высокую летальность. При низкой плотности мышей вспышки быстро затухают и, следовательно, имеют короткую продолжительность. При средней плотности эпизоотии медленно прожигают колонию, тогда как при высокой плотности котерии настолько хорошо связаны, что чума быстро распространяется по колонии.( C ) Индивидуальный R 0 . Белый цвет обозначает значения <1. ( D ) На основе Coterie R 0 ( R *) с тем же цветовым кодом.
Большой интерес представляет наблюдение, что чума способна персистировать в популяциях луговых собачек с низкой частотой в течение значительных периодов времени (например, энзоотические фазы продолжались более 1 года примерно в 25% прогонов модели). То есть наши результаты показывают, что чума может сохраняться в популяциях высоко восприимчивого хозяина в течение длительного времени и что нет необходимости ссылаться на существование альтернативных видов хозяев для объяснения межэпизоотической динамики чумы.
То есть наши результаты показывают, что чума может сохраняться в популяциях высоко восприимчивого хозяина в течение длительного времени и что нет необходимости ссылаться на существование альтернативных видов хозяев для объяснения межэпизоотической динамики чумы.
При включении в модель мышей-кузнечиков вероятность возникновения эпизоотии чумы возрастала. При низкой плотности мышей-кузнечиков активность чумы часто исчезала (поскольку мыши-кузнечики были настолько редки, что их территории не совпадали с котериями луговых собачек, содержащими заразные популяции блох), но низкая плотность мышей иногда вызывала массовое вымирание популяции луговых собачек. . Однако при более высокой плотности мышей-кузнечиков эпизоотии чумы, приводящие к вымиранию колонии луговых собачек, были практически гарантированы, хотя продолжительность эпизоотии могла достигать 4 лет (рис.1 В ). Результаты показывают четкое пороговое явление между численностью мышей и вспышками чумы среди луговых собачек: увеличение плотности мышей на несколько процентов приводит к резкому увеличению общего числа инфицированных луговых собачек. При низкой плотности мыши не способны распространять чуму по всей колонии, но после того, как плотность мышей превышает пороговое значение, частым исходом являются крупные эпизоотии у луговых собачек.
При низкой плотности мыши не способны распространять чуму по всей колонии, но после того, как плотность мышей превышает пороговое значение, частым исходом являются крупные эпизоотии у луговых собачек.
Чтобы лучше понять роль численности кузнечиковых мышей в динамике заболевания у основного хозяина луговых собачек, мы адаптировали модель, чтобы полностью предотвратить рассредоточение луговых собачек и аннексию брошенных группок соседними луговыми собачками.В результате чума распространяется исключительно через кузнечиков, а эпизоотии у основного хозяина вызываются исключительно альтернативным хозяином; следовательно, мы можем исследовать динамику заболевания с точки зрения сети. Порог перколяции возникает, когда вершины в сети соединены таким образом, что образуют гигантский компонент (т. е. связный подграф, содержащий большинство вершин в сети) (19). В контексте инфекционных заболеваний вершина представляет собой хозяина (или группу хозяев), а ребро представляет собой контакт с возможностью передачи болезни между двумя группами хозяев. Ниже порога перколяции не существует гигантского компонента, и первоначальная инфекция случайного хозяина не может распространиться на остальную часть популяции хозяина. В нашей модели чумы вершины представляют котерии луговых собачек, а ребро существует между двумя вершинами всякий раз, когда мышь-кузнечик перемещается из своей базовой котерии в другую котерию. Мы рассчитываем количество компонентов (одновершинные компоненты исключаются, поскольку они не имеют значения для распространения болезни) и средний размер компонентов.Начиная с очень низкой плотности мышей, увеличение плотности мыши сначала приводит к увеличению количества компонентов (поскольку каждая мышь, вероятно, создает новый компонент) (рис. 2). При определенной пороговой плотности количество компонентов начинает уменьшаться по мере увеличения плотности мыши просто потому, что дополнительные мыши вряд ли создадут новые компоненты; вместо этого они будут подключаться к уже существующим компонентам и потенциально даже соединять ранее отключенные компоненты.
Ниже порога перколяции не существует гигантского компонента, и первоначальная инфекция случайного хозяина не может распространиться на остальную часть популяции хозяина. В нашей модели чумы вершины представляют котерии луговых собачек, а ребро существует между двумя вершинами всякий раз, когда мышь-кузнечик перемещается из своей базовой котерии в другую котерию. Мы рассчитываем количество компонентов (одновершинные компоненты исключаются, поскольку они не имеют значения для распространения болезни) и средний размер компонентов.Начиная с очень низкой плотности мышей, увеличение плотности мыши сначала приводит к увеличению количества компонентов (поскольку каждая мышь, вероятно, создает новый компонент) (рис. 2). При определенной пороговой плотности количество компонентов начинает уменьшаться по мере увеличения плотности мыши просто потому, что дополнительные мыши вряд ли создадут новые компоненты; вместо этого они будут подключаться к уже существующим компонентам и потенциально даже соединять ранее отключенные компоненты. По мере добавления большего количества мышей порог просачивания пересекается (т.т. е. большинство вершин теперь связаны и образуют гигантский компонент), а среднее число компонентов быстро приближается к 1. В нашей модели чумы это означает, что почти все котерии теперь связаны движением мыши, и, таким образом, чума может распространяться и заражают большую часть популяции луговых собачек. Действительно, доля зараженной популяции луговых собачек резко возрастает по мере уменьшения количества компонентов, а среднее количество компонентов является надежным предиктором размера эпизоотии (ясно, что ни одна эпизоотия не может превышать количество хосты, связанные рассредоточением мыши, и, таким образом, пунктирная линия на рис.2 указан максимально возможный размер эпизоотии). То есть мы показываем, что поведение порога численности мышей представляет собой явление перколяции: при низкой плотности мыши могут распространять чуму только локально, но при более высокой плотности территории мышей перекрываются в достаточной степени, чтобы чума могла распространяться по всей колонии (рис.
По мере добавления большего количества мышей порог просачивания пересекается (т.т. е. большинство вершин теперь связаны и образуют гигантский компонент), а среднее число компонентов быстро приближается к 1. В нашей модели чумы это означает, что почти все котерии теперь связаны движением мыши, и, таким образом, чума может распространяться и заражают большую часть популяции луговых собачек. Действительно, доля зараженной популяции луговых собачек резко возрастает по мере уменьшения количества компонентов, а среднее количество компонентов является надежным предиктором размера эпизоотии (ясно, что ни одна эпизоотия не может превышать количество хосты, связанные рассредоточением мыши, и, таким образом, пунктирная линия на рис.2 указан максимально возможный размер эпизоотии). То есть мы показываем, что поведение порога численности мышей представляет собой явление перколяции: при низкой плотности мыши могут распространять чуму только локально, но при более высокой плотности территории мышей перекрываются в достаточной степени, чтобы чума могла распространяться по всей колонии (рис. 2).
2).
Компоненты сети и эпизоотические пороги. Серые столбцы показывают количество компонентов в колонии луговых собачек [т.е. в сети котерий (узлов), соединенных расселением мышей (ребра)] в зависимости от плотности мыши.Черная сплошная линия представляет среднюю долю зараженной популяции луговых собачек. Пунктирная линия представляет средний размер компонента графа (измеряемый как количество связанных котерий), выраженный как доля от общего количества котерий. Средний размер эпизоотии зависит от среднего размера компонента графа, поскольку любая вспышка ограничена размером компонента, в котором происходит первоначальная инфекция. Кривая вспышки немного смещена вниз относительно кривой компонента из-за стохастической передачи болезни.(Параметры установлены на значения по умолчанию, указанные в описании модели; j P = 0,6).
Способность предсказывать пороги заболеваемости традиционно основывалась на концепции R o , числе вторичных случаев, вызванных первым инфицированным человеком в полностью восприимчивой популяции. Однако было показано, что стандартный порог инвазии R o = 1 (выше которого болезнь может проникнуть) является плохим предиктором вспышек болезни в популяциях, которые подразделяются на группы с ограниченным перемещением между группами.В таких популяциях R *, эквивалентный на уровне группы R или (20), является лучшим предиктором инвазии болезни (21–23). Совсем недавно было показано, что порог проникновения возникает в популяциях больших песчанок в Центральной Азии, где вспышки болезней, по-видимому, происходят только при превышении определенного порога численности хозяев (24). Наша модель предполагает, что порог перколяции вторичного хозяина предсказывает эпизоотию чумы у луговых собачек (рис. 2). Чтобы показать, что этот порог перколяции отличается от порогов R o и R *, мы измерили индивидуальные и групповые R o (т.э., Р *). Индивидуальные R o – это среднее число вторичных инфекций луговых собачек, вызванных первой инфицированной луговой собачкой в изначально восприимчивой популяции.
Однако было показано, что стандартный порог инвазии R o = 1 (выше которого болезнь может проникнуть) является плохим предиктором вспышек болезни в популяциях, которые подразделяются на группы с ограниченным перемещением между группами.В таких популяциях R *, эквивалентный на уровне группы R или (20), является лучшим предиктором инвазии болезни (21–23). Совсем недавно было показано, что порог проникновения возникает в популяциях больших песчанок в Центральной Азии, где вспышки болезней, по-видимому, происходят только при превышении определенного порога численности хозяев (24). Наша модель предполагает, что порог перколяции вторичного хозяина предсказывает эпизоотию чумы у луговых собачек (рис. 2). Чтобы показать, что этот порог перколяции отличается от порогов R o и R *, мы измерили индивидуальные и групповые R o (т.э., Р *). Индивидуальные R o – это среднее число вторичных инфекций луговых собачек, вызванных первой инфицированной луговой собачкой в изначально восприимчивой популяции. Coterie-based R o ( R *) — это среднее количество групп, в которых хотя бы одна инфекция луговых собачек была вызвана луговыми собачками в первоначально инфицированной группе.
Coterie-based R o ( R *) — это среднее количество групп, в которых хотя бы одна инфекция луговых собачек была вызвана луговыми собачками в первоначально инфицированной группе.
R o не удалось точно предсказать вспышки чумы, потому что, хотя чума легко передается между луговыми собачками внутри группы, распространение болезни между группами в значительной степени зависит от мышей-кузнечиков и, таким образом, не зависит от R o ( Инжир.1 С ). R * является лучшим предиктором вспышек чумы, поскольку учитывает распространение болезни между группами. Тем не менее, это верно только в том случае, если плотность мышей достаточно высока (рис. 1 D ), потому что только тогда большинство котерий соединяются движением мыши, образуя гигантский компонент (рис. 2). При низкой плотности мышей группы котерий образуют множество изолированных небольших компонентов, и хотя R * правильно предсказывает передачу внутри этих компонентов, крупномасштабных вспышек произойти не может.
Недавние данные полевых исследований подтверждают описание нашей моделью экологии чумы в популяциях луговых собачек. Высокая численность мышей-кузнечиков была связана с повышенной вероятностью гибели луговых собачек, вызванной чумой, в течение следующего года (17). Кроме того, растущее число мышей-кузнечиков заражается блохами луговых собачек за несколько месяцев до того, как действительно наблюдается крупномасштабная гибель луговых собачек, что мы интерпретируем как свидетельство гибели семейных групп луговых собачек от чумы, хотя популяция луговых собачек снижается. трудно обнаружить (17). Зараженные Y. pestis блохи были выделены из колоний луговых собачек за несколько месяцев до эпизоотии (25). Кроме того, генетическая структура популяции Y. pestis в популяциях аризонских луговых собачек указывает на периоды очень быстрой передачи чумы, за которыми следовали периоды более медленной локализованной дифференциации (26).
Мы описали механизм активности чумы в одной колонии луговых собачек, который включает обострение передачи болезни среди популяции луговых собачек в сочетании с высокой плотностью мышей-кузнечиков. Характер вымирания колоний луговых собачек предполагает, что площадь колонии и судьба соседних колоний оказывают важное влияние на вероятность вымирания колонии (8). Эта закономерность предполагает, что перемещение чумы между колониями луговых собачек вызвано перемещением инфекционных животных или животных, переносящих инфекционных блох, и что динамика метапопуляции может иметь важное значение для объяснения экологии чумы в ландшафтном масштабе (1, 8, 26, 27). Тем не менее, наша модель и полевые наблюдения показывают, что, если чума достигнет колонии луговых собачек, местная численность кузнечиков окажет сильное влияние на вероятность появления Y.pestis переросла в вспышку, а не загадочно тлела в популяции луговых собачек. Численность мышей-кузнечиков колеблется со временем, а также варьируется в зависимости от ландшафта (17, 28), что позволяет предположить, что гибель луговых собачек является следствием совпадения динамики популяции мышей и перемещения Y. pestis (т.
Характер вымирания колоний луговых собачек предполагает, что площадь колонии и судьба соседних колоний оказывают важное влияние на вероятность вымирания колонии (8). Эта закономерность предполагает, что перемещение чумы между колониями луговых собачек вызвано перемещением инфекционных животных или животных, переносящих инфекционных блох, и что динамика метапопуляции может иметь важное значение для объяснения экологии чумы в ландшафтном масштабе (1, 8, 26, 27). Тем не менее, наша модель и полевые наблюдения показывают, что, если чума достигнет колонии луговых собачек, местная численность кузнечиков окажет сильное влияние на вероятность появления Y.pestis переросла в вспышку, а не загадочно тлела в популяции луговых собачек. Численность мышей-кузнечиков колеблется со временем, а также варьируется в зависимости от ландшафта (17, 28), что позволяет предположить, что гибель луговых собачек является следствием совпадения динамики популяции мышей и перемещения Y. pestis (т. е. одновременного взаимодействия перемещения болезни и хозяина). экология). Интересно, что наши результаты, основанные на модельной интерпретации эмпирических наблюдений, перекликаются с предыдущими теоретическими работами по изучению персистенции и вспышек заболеваний.
е. одновременного взаимодействия перемещения болезни и хозяина). экология). Интересно, что наши результаты, основанные на модельной интерпретации эмпирических наблюдений, перекликаются с предыдущими теоретическими работами по изучению персистенции и вспышек заболеваний.
Заключение
Таким образом, использование полевых данных для параметризации пространственно точной многохозяинной модели динамики чумы дало неожиданное понимание экологии болезни, которое позволяет предположить, что ( i ) чума способна сохраняться в колониях луговых собачек в течение длительных периодов времени при низких температурах. энзоотические уровни, просто перемещаясь последовательно между соседними семейными группами (котериями) и ( ii ) кузнечиковых мышей, даже при низких частотах усугубляют распространение чумы в колонии луговых собачек, вызывая крупные эпизоотии.Эти результаты дают представление о динамике чумы, то есть о том, что сложные взаимодействия с несколькими хозяевами вызывают вспышки болезни, но процесс распространения от энзоотических хозяев не обязательно имеет решающее значение для персистенции возбудителя между эпизоотиями. Действительно, склонность чумы к социальным роющим животным, сохраняющим территорию (например, луговые собачки, суслики, большие песчанки и т. д.) (1), может объяснить ее способность сохраняться в популяции хозяина в качестве высоколетального патогена.
Действительно, склонность чумы к социальным роющим животным, сохраняющим территорию (например, луговые собачки, суслики, большие песчанки и т. д.) (1), может объяснить ее способность сохраняться в популяции хозяина в качестве высоколетального патогена.
Наблюдение, что дополнительные виды могут сильно влиять на динамику болезни, особенно интересно, потому что оно иллюстрирует классический феномен перколяционного порога.Хотя важность перколяционных порогов в системах болезней была установлена ранее (24), ранее не сообщалось о пороговых значениях перколяции, в которых альтернативные хозяева являются ключевым элементом, определяющим вспышки болезни у первичного хозяина. Многие системы болезней включают несколько хозяев, инфицированных универсальными патогенами (например, вирус чумы плотоядных собак Серенгети и болезнь Лайма у мелких млекопитающих) (29, 30), и, кроме того, включают виды, занимающие более или менее дискретные ареалы обитания, которые индуцируют соответствующую пространственную структуру. к передаче болезни.Следовательно, понимание динамики перколяции с участием альтернативных хозяев может иметь решающее значение для понимания сложной динамики болезней диких животных и зоонозов.
к передаче болезни.Следовательно, понимание динамики перколяции с участием альтернативных хозяев может иметь решающее значение для понимания сложной динамики болезней диких животных и зоонозов.
Материалы и методы
Мы разработали простую вычислительную модель для моделирования динамики чумы в колониях луговых собачек, преимущественно используя данные нашего исследовательского полигона в Национальном пастбище Пауни (Pawnee National Grasslands, PNG), север Колорадо (17). В Папуа-Новой Гвинее колонии луговых собачек имеют размер до 250 га, а плотность луговых собачек по консервативным расчетам составляет 22 га −1 (31), что позволяет предположить, что большие колонии населены более чем 5500 луговыми собачками.Территории Coterie (семейных групп) обычно являются смежными (18) и на северо-западе Колорадо обычно занимают около 0,2 га (диапазон = 0,12–0,61 га) (32). Семьи луговых собачек защищают территорию своего кружка от соседних луговых собачек, но в случае гибели всех членов кружка луговые собачки обычно присоединяются к одной соседней семье (18). Распространение луговых собачек ограничено 10-недельным окном каждый год (33).
Распространение луговых собачек ограничено 10-недельным окном каждый год (33).
Мыши-кузнечики многочисленны в колониях луговых собачек (28) с плотностью 0–10 га −1 или эквивалентно до 2 мышей на группу луговых собачек.Мыши-кузнечики с радиоотслеживанием в колониях луговых собачек имели средние участки обитания 3,84 га (SD = 1,64) при наблюдении в течение 7-дневного периода, что предполагает, что мыши пересекают и взаимодействуют по крайней мере с тремя группами за ночь.
Луговые собачки в основном заражаются блохой O. hirsuta (25). После заражения чумой луговые собачки погибают примерно через 6 дней (12, 34), а блохи впоследствии ищут новых хозяев. Умирающие луговые собачки заражают большую часть своих блох бактерией чумы: 18/22 (или 82%) O.hirsuta , обнаруженные на мертвых луговых собачках, оказались чумно-позитивными, и блохи, вероятно, заражаются только на терминальных стадиях инфекции хозяина Y. pestis (35, 36). Мыши-кузнечики обычно редко являются хозяевами O.![]() hirsuta , но во время эпизоотий чумы 81% мышей-кузнечиков могут быть заражены этим видом блох (17).
hirsuta , но во время эпизоотий чумы 81% мышей-кузнечиков могут быть заражены этим видом блох (17).
Мы смоделировали динамику чумы в одной большой колонии луговых собачек, организованной в виде квадратной сетки из 33 × 33 участков (всего = 1089), где каждый участок изначально занят одной группой луговых собачек, состоящей из пяти особей и обеспечивающей первоначальный размер популяции 5445 луговых собачек.Мыши-кузнечики были распределены по каждому участку в рамках испытания Бернулли с вероятностью p M (т. е. для p M = 0 динамика чумы моделируется без присутствия мышей) и были распределены по трем случайным группам в поблизости от их базового кружка.
Мы моделируем одну большую колонию как пространственную регулярную сеть, где каждая вершина представляет группу, изначально населенную одной семьей из пяти луговых собачек. Моделирование выполняется для 1825 временных шагов, где каждый временной шаг представляет собой день (чтобы учесть ежедневные различия в вероятности передачи чумы), а моделирование аппроксимирует динамику чумы за 5 лет. В случайный день в течение первых 365 дней моделирования случайно выбранная особь луговой собачки заражается чумой. На каждом временном шаге происходят следующие процессы: ( i ) воздействие на луговых собачек инфекционных блох, спрыгивающих с туш убитых чумой луговых собачек, ( ii ) передача Y. pestis от блох луговым собачкам, ( iii ) воздействие на луговых собачек блох, спрыгивающих с мышей, и ( iv ) воздействие на мышей инфекционных блох и Y.инфекция pestis . Более подробную информацию можно найти в SI Materials and Methods .
В случайный день в течение первых 365 дней моделирования случайно выбранная особь луговой собачки заражается чумой. На каждом временном шаге происходят следующие процессы: ( i ) воздействие на луговых собачек инфекционных блох, спрыгивающих с туш убитых чумой луговых собачек, ( ii ) передача Y. pestis от блох луговым собачкам, ( iii ) воздействие на луговых собачек блох, спрыгивающих с мышей, и ( iv ) воздействие на мышей инфекционных блох и Y.инфекция pestis . Более подробную информацию можно найти в SI Materials and Methods .
Чтобы понять закономерности динамики чумы, мы исследовали пространство параметров, запустив 100 симуляций для каждой комбинации параметров. (Для нескольких комбинаций тестовых параметров мы выполнили 1000 симуляций, и наблюдаемые закономерности оставались устойчивыми). Мы использовали следующие параметры: j P , вероятность того, что блохи набросятся на хозяев, бросивших убитых чумой луговых собачек, варьировалась от 0. от 1 до 1,0, частота мыши варьировалась от 0 до 1,0, а рассредоточение луговых собачек по случайным территориям группы варьировалось от 0% до 20%. Мы измерили три выхода: общее количество инфицированных луговых собачек, максимальную ежедневную частоту атак (количество луговых собачек, впервые инфицированных в течение 1 дня) и продолжительность активности чумы (продолжительность времени между первым заражением луговой собачки и последняя инфекция луговой собачки).
от 1 до 1,0, частота мыши варьировалась от 0 до 1,0, а рассредоточение луговых собачек по случайным территориям группы варьировалось от 0% до 20%. Мы измерили три выхода: общее количество инфицированных луговых собачек, максимальную ежедневную частоту атак (количество луговых собачек, впервые инфицированных в течение 1 дня) и продолжительность активности чумы (продолжительность времени между первым заражением луговой собачки и последняя инфекция луговой собачки).
Объяснение концовки «Сила собаки»: судьбы Фила и Питера
Предупреждение о спойлерах: в следующем интервью обсуждаются ключевые детали «Силы собаки» от Netflix, включая концовку.Если вы еще не видели фильм, вернитесь, когда посмотрите, и ознакомьтесь с нашим обзором, профилем режиссера Джейн Кэмпион и интервью с партнершей по фильму Кирстен Данст.
Через пять минут после начала «Собачьей силы» мы встречаем Питера (Коди Смит-МакФи), хилого молодого человека, сидящего в одиночестве в своей комнате и кропотливо мастерящего бумажные цветы, которые станут украшением стола в гостиной. гостиница-ресторан, которой владеет его мать Роуз (Кирстен Данст).
гостиница-ресторан, которой владеет его мать Роуз (Кирстен Данст).
Вечером того же дня эти цветы превратятся в пепел, сожженный запугивающим владельцем ранчо Филом (Бенедикт Камбербэтч), первым в серии угрожающих действий, направленных против Питера и его матери, угроза, которая придает вестерну коммерческую пьесу Джейн Кэмпион. , почти невыносимое напряжение — которое Смит-Макфи остро ощутил при первом прочтении сценария.
«Мне все время было жалко Питера, — говорит 25-летний Смит-Макфи во время недавней беседы в ресторане Санта-Моники. «У меня было так много суждений о том, как он себя вел. В конце концов эти суждения укрепились, и у меня просто возникло ощущение надвигающейся на него гибели».
А потом мы понимаем, что, возможно, мы плохо видели Питера.
Смит-Макфи с нами. Дойдя до конца своего первого прохода по сценарию, он пролистал несколько страниц назад, чтобы убедиться, что ничего не пропустил.
«Я ничего не пропустил», — улыбаясь, говорит энергичный австралийский актер.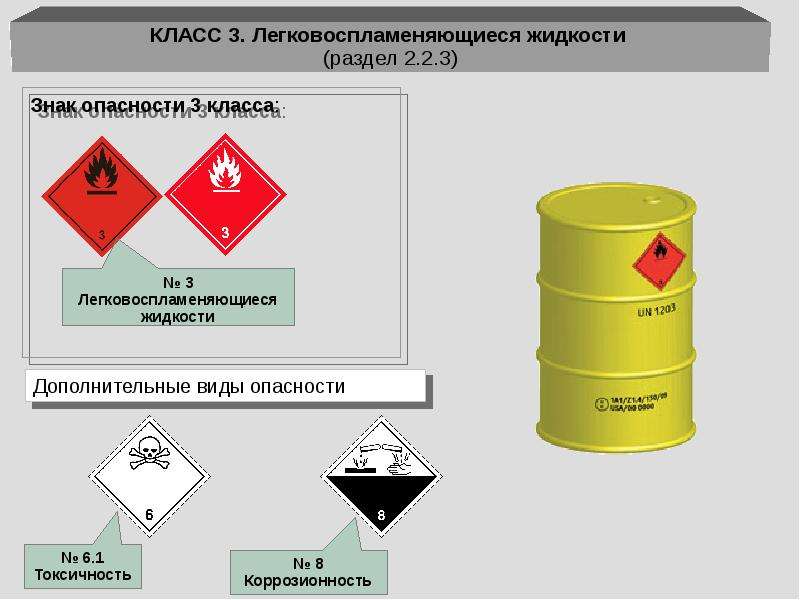
Если вы видели «Сила собаки» — и, опять же, если нет, добавьте эту страницу в закладки для дальнейшего чтения — вам может быть интересно, не пропустили ли вы что-нибудь. Смит-МакФи, новоиспеченный победитель конкурса актеров второго плана в Нью-Йоркском кружке кинокритиков, здесь, чтобы ответить на все ваши вопросы, от причин, по которым Питер выбрал обувь, до удивительного финала фильма.
Когда Питер решает убить Фила?
Это неоднозначно, так что это просто я… но я думаю, что это когда он видит, как он сжигает цветы.
Все восходит к цветам? Я думал, что это было вызвано тем, что его мать впала в алкоголизм из-за жестокости Фила.
Конечно. Но Фил определенно попал в поле зрения Питера с той первой ночи. Это красивая метафора. Горящие розы Фила, что он собирается сделать с матерью Питера, уничтожив ее. Но я думаю, что Питера также расстроило то, что Фил довел его почти до слез во время первой встречи.Он не знал, что его можно так легко ранить.
Да, Питер выходит на улицу, и мы видим, как он крутит хулахуп, который я сначала рассматривал как приспособление, вроде тех спиннеров, которые были так популярны несколько лет назад.
Похоже, он пытается успокоить свою тревогу. Такое ощущение, что он потерял контроль. Но все совсем наоборот. Он управляет. Он абсолютно все контролирует.
То же самое, когда он проводит пальцами по зубьям расчески, что мы видим во время явного стресса.Первоначальное впечатление, что он пытается справиться… или досадить матери. Или оба!
Знаешь, то, что я делал с расческой, успокаивало меня, когда я играл с ним. Но я думаю, что для Питера это способ управлять своими мыслями и эмоциями. Это физическое проявление того, как работает его разум, порождая различные мысли и планы. То же и с хулахупом. Вернитесь и посмотрите его, и я не думаю, что вы когда-нибудь увидите, как кто-то крутит обруч с таким гневом.
ОК. Итак, вы думаете, что Питер всегда планировал убить Фила, и когда он узнает о телятах, умирающих от сибирской язвы, этот лакомый кусочек порождает идею претворить план в жизнь?
Избавление коровы от сибирской язвы, как и убийство кролика в его комнате, отчасти помогает ему стать врачом. Но Питер импровизатор. Он плывет по течению. И когда он видит, что Фил порезал ему руку, это наводит его на мысль заразить сыромятную кожу и заразить его.
Но Питер импровизатор. Он плывет по течению. И когда он видит, что Фил порезал ему руку, это наводит его на мысль заразить сыромятную кожу и заразить его.
Питер постоянно оценивает, ожидая идеального момента для удара. Я думаю, это свойственно многим животным, но для Питера я выбрал лису, чтобы слиться с его персонажем. Они такие легкие на ногах, красивые и выглядят нежными и милыми. И все же они могут на цыпочках подойти к дыре и прыгнуть прямо туда, и вы пропали.
Бенедикт Камбербэтч в роли Фила в фильме «Сила собаки».
(Kirsty Griffin/ Netflix)
Что приводит нас к сцене в сарае, где Питер заражает Фила сибирской язвой. Это такой тонкий танец между этими двумя мужчинами, и осмелюсь сказать…
Тебе было жаль Фила? Я рядом с тобой. Стены Фила, стены, которые так долго застывали, исчезли, а его маска снята. Узнаем правду о его любви к Бронко Генри. И, да, он, возможно, был подготовлен, но я думаю, что то, что у него было с Бронко Генри, было подлинным. Он был влюблен. И ему приходилось скрывать это от мира. Тогда как Питер… Фил завидует Питеру, потому что он может быть полностью самим собой. Интересно, как они меняются ролями. Питер становится более мрачной и жестокой фигурой, а Фил — более невинным ребенком с большой травмой. Когда Питер держит зараженную веревку в перчатках, я думаю, он думает о Филе. И это больно.
Он был влюблен. И ему приходилось скрывать это от мира. Тогда как Питер… Фил завидует Питеру, потому что он может быть полностью самим собой. Интересно, как они меняются ролями. Питер становится более мрачной и жестокой фигурой, а Фил — более невинным ребенком с большой травмой. Когда Питер держит зараженную веревку в перчатках, я думаю, он думает о Филе. И это больно.
Как вы думаете, Питер когда-нибудь думал отказаться от своего плана и пощадить Фила?
№То же самое было и с кроликом, которого он убил, чтобы препарировать его. Он собирается делать то, что должен. И хотя для Питера это может быть его первое взаимодействие с близостью или любовью к другому человеку, он готов пожертвовать этим ради своей матери.
Вы упомянули о том, что Фил завидует Питеру, потому что он так хладнокровен, что напомнило мне белые туфли Питера. Они выглядят такими неуместными на ранчо… что, опять же, немного неверно, не так ли?
Да! Туфли символизируют уверенность Петра и его непоколебимый дух перед судом.